LAG3:腫瘤免疫治療的下一個風(fēng)口
日期:2025-07-18 11:41:53
1. LAG3的背景介紹
淋巴細(xì)胞活化基因3(LAG3,CD223)是一種重要的免疫檢查點分子,于1990年首次被克隆,其基因與CD4基因同源性約20%,編碼含498個氨基酸的跨膜蛋白,包含四個免疫球蛋白超家族(IgSF)結(jié)構(gòu)域(D1-D4),獨特的胞質(zhì)尾含有 “KIEELE” 基序和谷氨酸-脯氨酸(EP)重復(fù)序列,這些結(jié)構(gòu)是其發(fā)揮免疫調(diào)節(jié)功能的關(guān)鍵 [1]。
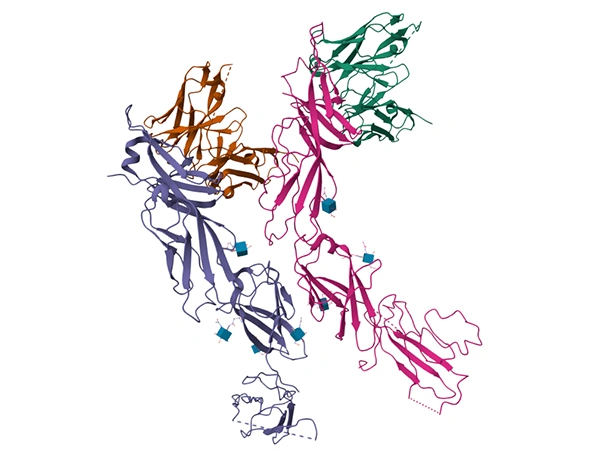
Human LAG3的結(jié)構(gòu)(圖源:PDB 7TZG)
LAG3主要表達(dá)于活化的T細(xì)胞(CD4?、CD8?)、調(diào)節(jié)性T細(xì)胞(Tregs)、NK細(xì)胞、B細(xì)胞及plasmacytoid樹突狀細(xì)胞(pDCs)等免疫細(xì)胞表面 [2,3]。在腫瘤微環(huán)境中,LAG3常與PD1等其他抑制性受體共表達(dá)于耗竭的T細(xì)胞上,通過抑制T細(xì)胞增殖、細(xì)胞因子分泌及細(xì)胞毒性功能,參與腫瘤免疫逃逸 [1,3]。
2. LAG3的研究機制
2.1 與 TCR-CD3 復(fù)合物的相互作用
LAG3可與T細(xì)胞受體(TCR-CD3 復(fù)合物持續(xù)關(guān)聯(lián),在T細(xì)胞活化后遷移至免疫突觸(IS)。其胞質(zhì)尾的EP重復(fù)序列通過降低免疫突觸局部pH,破壞CD4/CD8共受體與酪氨酸激酶Lck之間的鋅離子(Zn2?)結(jié)合,導(dǎo)致共受體-Lck復(fù)合物解離,最終抑制TCR下游信號(如ZAP70、ERK、AKT磷酸化),限制T細(xì)胞活化。這一過程不依賴其經(jīng)典配體MHCⅡ類分子,是LAG3獨立抑制T細(xì)胞功能的核心機制 [3]。
2.2 配體結(jié)合與二聚化機制
LAG3的主要配體包括MHCⅡ類分子、纖維蛋白原樣蛋白1(FGL1)、肝竇內(nèi)皮細(xì)胞凝集素(LSECtin)等 [2]。其D1結(jié)構(gòu)域的Loop 2區(qū)域(Gly1??-Pro11?)是配體結(jié)合的關(guān)鍵界面,與MHCⅡ類分子、FGL1的結(jié)合均依賴該區(qū)域 [4]。此外,LAG3通過D2結(jié)構(gòu)域形成同源二聚體, murineLAG3二聚體由Trp1??與相鄰protomer的疏水口袋相互作用介導(dǎo),人類LAG3二聚體則依賴Phe22?、Trp1??等殘基,二聚化可增強其與配體的結(jié)合能力。
2.3 與其他免疫檢查點的協(xié)同作用
LAG3與PD1在T細(xì)胞耗竭中存在協(xié)同效應(yīng)。在慢性感染和腫瘤模型中,單獨阻斷LAG3或PD1對T細(xì)胞功能的恢復(fù)有限,而聯(lián)合阻斷可顯著逆轉(zhuǎn)T細(xì)胞耗竭,增強細(xì)胞因子分泌和增殖能力[1]。這種協(xié)同可能與二者分別通過不同機制抑制TCR信號有關(guān):LAG3主要干擾共受體-Lck相互作用,PD1則通過SHP-1/2抑制下游信號 [1,3]。
3. LAG3相關(guān)信號通路
LAG3主要通過以下途徑抑制免疫應(yīng)答:
- TCR信號抑制:通過EP重復(fù)序列降低局部pH,破壞CD4/CD8與Lck的結(jié)合,減少ZAP70、PLCγ等分子的磷酸化,抑制鈣內(nèi)流和NFAT等轉(zhuǎn)錄因子激活 [3];
- Treg細(xì)胞功能增強:LAG3在Treg細(xì)胞上高表達(dá),通過與MHCⅡ類分子結(jié)合,增強Treg的免疫抑制功能,抑制效應(yīng)T細(xì)胞增殖 [2];
- 抗原呈遞細(xì)胞(APC)調(diào)節(jié):LAG3與DC表面的MHCⅡ類分子結(jié)合,可抑制DC成熟,減少IL-12等促炎細(xì)胞因子分泌,促進IL-10等抑炎因子產(chǎn)生 [1,2]。
4. LAG3相關(guān)疾病
IL2的異常調(diào)控會導(dǎo)致免疫失衡、信號通路紊亂等,與多種疾病的發(fā)生發(fā)展密切相關(guān):
4.1 腫瘤?
4.1.1 葡萄膜黑色素瘤(UM)
在43例UM組織樣本中,25例(58.1%)檢測到LAG3表達(dá),且LAG3同時存在于腫瘤細(xì)胞和腫瘤浸潤淋巴細(xì)胞(TILs)中。具有組織病理學(xué)高危因素(如大細(xì)胞型、壞死等)的UM病例中,LAG3基因表達(dá)顯著上調(diào);LAG3陽性TILs及mRNA高表達(dá)的患者無病生存率(DFS)顯著降低(p<0.0001),提示LAG3可作為UM高危患者的預(yù)后生物標(biāo)志物。其機制可能與LAG3抑制TILs的抗腫瘤活性有關(guān),導(dǎo)致腫瘤細(xì)胞逃逸免疫監(jiān)視 [5]。
4.1.2 胃腸胰神經(jīng)內(nèi)分泌腫瘤(GEP-NETs)
對61例GEP-NETs患者的分析顯示,所有患者均未檢測到LAG3陽性表達(dá)(0/61),但PD-L1、MSI-H等其他免疫標(biāo)志物存在表達(dá)差異:PD-L1在G3級腫瘤中陽性率最高(50%),MSI-H在G1級中占80%,且二者在局部病變中更常見。盡管LAG3在該腫瘤中未顯陽性,但其與其他免疫檢查點的協(xié)同缺失可能參與GEP-NETs的免疫逃逸,具體機制需進一步研究 [6]。
4.1.3 乳腺癌
乳腺癌組織中,LAG3?上皮內(nèi)腫瘤浸潤淋巴細(xì)胞(iTILs)的存在與改善的乳腺癌特異性生存率(BCSS)相關(guān)(HR 0.71;95% CI 0.56–0.90)。約53%的PD-L1?腫瘤和61%的PD-1?腫瘤中存在LAG3? iTILs,且LAG3? CD8? iTILs共浸潤時,BCSS更優(yōu)。這提示LAG3? iTILs可能反映機體抗腫瘤免疫活性,其存在預(yù)示更有效的免疫應(yīng)答 [7]。
4.1.4 非小細(xì)胞肺癌(NSCLC)
LAG3在NSCLC腫瘤微環(huán)境的TILs上高表達(dá),且與腫瘤分期相關(guān):Ⅲ期患者腫瘤微循環(huán)中LAG3?細(xì)胞比例高于Ⅳ期,同時LAG3表達(dá)與PD-L1表達(dá)呈正相關(guān)。高LAG3表達(dá)的患者對PD-1抑制劑的應(yīng)答率較低,無進展生存期(PFS)縮短,提示LAG3可能是NSCLC免疫治療耐藥的標(biāo)志物。機制可能與LAG3與PD1協(xié)同抑制T細(xì)胞功能,削弱免疫治療效果有關(guān) [8]。
4.1.5 胃癌
在接受nivolumab單藥治療的晚期胃癌患者中,LAG3? TILs的存在與治療應(yīng)答率降低相關(guān),且LAG3 mRNA水平高的患者總生存期(OS)顯著縮短。此外,LAG3與腫瘤突變負(fù)荷(TMB)呈負(fù)相關(guān),提示LAG3高表達(dá)可能通過抑制T細(xì)胞浸潤,降低免疫治療敏感性 [9,10]。
4.1.6 結(jié)腸癌
在錯配修復(fù)缺陷(MMR-deficient)的局部晚期結(jié)腸癌中,新輔助nivolumab聯(lián)合relatlimab(抗LAG3)治療可誘導(dǎo)顯著的病理完全緩解(pCR),且腫瘤微環(huán)境中CD8? T細(xì)胞浸潤增加,LAG3?細(xì)胞比例降低。這表明LAG3在結(jié)腸癌中通過抑制細(xì)胞毒性T細(xì)胞功能促進腫瘤進展,阻斷LAG3可恢復(fù)免疫應(yīng)答 [11]。
4.2 自身免疫疾病
LAG3缺陷或功能異常與自身免疫疾病易感性增加相關(guān)。在1型糖尿病模型中,Lag3?/?小鼠胰島浸潤的CD4?、CD8? T細(xì)胞數(shù)量顯著增加,疾病進展加速;在類風(fēng)濕關(guān)節(jié)炎(RA)患者中,LAG3? Treg細(xì)胞比例降低,且與疾病活動度呈負(fù)相關(guān),提示LAG3? Treg的免疫抑制功能受損可能加劇關(guān)節(jié)炎癥。機制主要與LAG3對效應(yīng)T細(xì)胞的抑制減弱、Treg功能下降有關(guān),導(dǎo)致自身反應(yīng)性T細(xì)胞過度活化 [2]。
4.3 慢性感染
在慢性乙型肝炎病毒(HBV)感染中,LAG3?紅細(xì)胞祖細(xì)胞通過分泌TGF-β,抑制HBsAg血清清除,且LAG3表達(dá)水平與HBsAg滴度呈正相關(guān)。其機制為LAG3?細(xì)胞通過LAG3與T細(xì)胞相互作用,同時釋放TGF-β抑制T細(xì)胞對HBV的清除能力,延緩病毒清除 [12]。
5. LAG3 靶向藥物研究進展
LAG3(淋巴細(xì)胞激活基因3)靶向藥物的研究進展迅速,目前已有多種藥物進入臨床試驗階段。其中,BMS-986016(relatlimab)作為首個LAG3阻斷劑,已開展多項Ⅰ至Ⅲ期臨床試驗,顯示出良好的耐受性和抗腫瘤活性。其他抗LAG3單克隆抗體如LAG525、TSR-033、INCAGN02385等也處于臨床試驗階段,初步數(shù)據(jù)表明這些藥物具有良好的安全性和潛在的抗腫瘤效果。同時,雙特異性抗體如MGD013(同時靶向LAG3和PD-1)也顯示出良好的耐受性和抗腫瘤活性。LAG3靶向藥物在單藥或聯(lián)合治療中均顯示出潛力,部分在研藥物管線列舉如下:
| 藥物 | 藥物類型 | 在研適應(yīng)癥(疾病名) | 在研機構(gòu) | 最高研發(fā)階段 |
|---|---|---|---|---|
| 納武利尤單抗/瑞拉利單抗 | 單克隆抗體 | 轉(zhuǎn)移性黑色素瘤 | 不可切除的黑色素瘤 | 黑色素瘤 | 腫瘤 | 轉(zhuǎn)移性結(jié)直腸癌 | 轉(zhuǎn)移性微衛(wèi)星穩(wěn)定結(jié)直腸癌等 | Bristol Myers Squibb Co. | Bristol-Myers Squibb Australia Pty Ltd. | Bristol-Myers Squibb Pharma EEIG | 百時美施貴寶(中國)投資有限公司 | Ono Pharmaceutical Co., Ltd. | 中美上海施貴寶制藥有限公司 | 批準(zhǔn)上市 |
| Fianlimab/Cemiplimab | 單克隆抗體 | 不可切除的黑色素瘤 | 頭頸部鱗狀細(xì)胞癌 | Regeneron Pharmaceuticals, Inc. | 臨床3期 |
| 重組人源淋巴細(xì)胞激活基因-3(hLAG-3)融合蛋白(Immutep) | Fc融合蛋白 | 轉(zhuǎn)移性非小細(xì)胞肺癌 | 轉(zhuǎn)移性HER2陰性乳腺癌 | HER2陽性轉(zhuǎn)移性乳腺癌 | 軟組織肉瘤 | 晚期頭頸部鱗狀細(xì)胞癌 | 乳腺癌 | 非小細(xì)胞肺癌 | 頭頸部鱗狀細(xì)胞癌等 | Immutep SAS | 泰州億佰康醫(yī)藥科技有限公司 | Merck Sharp & Dohme LLC | Immutep Ltd. | 泰州億騰景昂藥業(yè)股份有限公司 | 臨床3期 |
| Alcestobart | 單克隆抗體 | 腫瘤 | 轉(zhuǎn)移性食管鱗狀細(xì)胞癌 | 食管鱗狀細(xì)胞癌 | 實體瘤 | 結(jié)直腸癌 | 鼻咽癌 | 微衛(wèi)星穩(wěn)定型結(jié)直腸癌 | 晚期癌癥 | 晚期惡性實體瘤等 | 廣州百濟神州生物制藥有限公司 | 博謙生技股份有限公司 | 百濟神州(廣州)醫(yī)藥有限公司 | BeOne Medicines Ltd. | 南京維立志博生物科技股份有限公司 | 臨床3期 |
| 瑞拉利單抗 | 單克隆抗體 | 復(fù)發(fā)性非鱗狀非小細(xì)胞肺癌 | 不可切除的黑色素瘤 | 腎細(xì)胞癌 | 肝細(xì)胞癌 | 轉(zhuǎn)移性非小細(xì)胞肺癌等 | Bristol Myers Squibb Co. | 中美上海施貴寶制藥有限公司 | Ono Pharmaceutical Co., Ltd. | 臨床3期 |
| Favezelimab/Pembrolizumab | 單克隆抗體 | 大腸腺癌 | 轉(zhuǎn)移性結(jié)直腸癌 | 皮膚鱗狀細(xì)胞癌 | 錯配修復(fù)缺陷子宮內(nèi)膜癌 | 難治性經(jīng)典霍奇金淋巴瘤等 | 默沙東研發(fā)(中國)有限公司 | Merck Sharp & Dohme Corp. | Merck Sharp & Dohme LLC | Merck & Co., Inc. | 臨床3期 |
| Fianlimab | 單克隆抗體 | 黑色素瘤 | 轉(zhuǎn)移性黑色素瘤 | 晚期非小細(xì)胞肺癌 | 局部晚期非小細(xì)胞肺癌 | 肌層浸潤性膀胱癌等 | Regeneron Pharmaceuticals, Inc. | Memorial Sloan Kettering Cancer Center | Sanofi | 臨床3期 |
| 特泊利單抗 | 雙特異性抗體 | 胃食管交界處癌 | HER2陽性胃癌 | 轉(zhuǎn)移性 HER2 陽性胃食管結(jié)合部癌 | MacroGenics, Inc. | 臨床2/3期 |
| Nivolumab/Relatlimab/Hyaluronidase | 單克隆抗體 | 酶 | 轉(zhuǎn)移性黑色素瘤 | Bristol Myers Squibb Co. | 臨床2期 |
| FS-118 | 雙特異性抗體 | 非小細(xì)胞肺癌 | 彌漫性大B細(xì)胞淋巴瘤 | 頭頸部腫瘤 | 血液腫瘤 | 結(jié)腸癌 | Merck Serono SA | F-star Therapeutics, Inc. | invoX Pharma Ltd. | F-Star Delta Ltd. | F Star Co., Ltd. | 臨床2期 |
| GSK-2831781 | 單克隆抗體 | 潰瘍性結(jié)腸炎 | 銀屑病 | Immutep Ltd. | 臨床2期 |
| Tuparstobart | 單克隆抗體 | 轉(zhuǎn)移性Merkel細(xì)胞癌 | 頭頸部鱗狀細(xì)胞癌 | 轉(zhuǎn)移性黑色素瘤 | 不可切除的黑色素瘤 | Incyte Biosciences International SARL | Incyte Corp. | 臨床2期 |
| HLX-26 | 單克隆抗體 | 晚期肺非鱗狀非小細(xì)胞癌 | 晚期非小細(xì)胞肺癌 | 轉(zhuǎn)移性非小細(xì)胞肺癌 | 淋巴瘤 | 實體瘤 | 轉(zhuǎn)移性實體瘤 | 晚期惡性實體瘤 | 肝細(xì)胞癌 | 上海復(fù)宏漢霖生物技術(shù)股份有限公司 | 上海復(fù)星醫(yī)藥產(chǎn)業(yè)發(fā)展有限公司 | 上海復(fù)宏漢霖生物制藥有限公司 | 臨床2期 |
| DNV-3 | 單克隆抗體 | 黑色素瘤 | 食管鱗狀細(xì)胞癌 | 胃食管交界處癌 | 肝癌 | 淋巴瘤 | 小細(xì)胞肺癌等 | 浙江時邁藥業(yè)有限公司 | 臨床2期 |
| Tombestomig | 雙特異性抗體 | PD-L1陽性三陰性乳腺癌 | 腎細(xì)胞癌 | 局部晚期腎細(xì)胞癌 | 局部晚期尿路上皮癌 | 尿路上皮癌等 | Hoffmann-La Roche, Inc. | Roche Holding AG | F. Hoffmann-La Roche Ltd. | 羅氏(中國)投資有限公司 | 臨床2期 |
| Miptenalimab | 單克隆抗體 | 食管腺癌 | 晚期癌癥 | 晚期子宮內(nèi)膜癌 | 局部晚期不可切除的胃腺癌 | 黑色素瘤等 | Boehringer Ingelheim GmbH | 臨床2期 |
| Cugrastomig | 雙特異性抗體 | 大腸腺癌 | 非小細(xì)胞肺癌 | 非小細(xì)胞肺癌IIIA期 | 頭頸部鱗狀細(xì)胞癌等癌 | 肝轉(zhuǎn)移 | 黑色素瘤 | 不可切除的黑色素瘤 | 復(fù)發(fā)性經(jīng)典霍奇金淋巴瘤 | 中山康方生物醫(yī)藥有限公司 | 康方匯科(上海)生物有限公司 | 臨床1/2期 |
| Fanastomig | 雙特異性抗體 | 實體瘤 | 岸邁生物科技(蘇州)有限公司 | 臨床1/2期 |
| 重組全人抗LAG-3單克隆抗體(廣州譽衡生物科技/無錫藥明康德) | 單克隆抗體 | 晚期非小細(xì)胞肺癌 | 晚期惡性實體瘤 | 黑色素瘤 | 廣州譽衡生物科技有限公司 | 臨床1/2期 |
| Favezelimab | 單克隆抗體 | 血液腫瘤 | Merck Sharp & Dohme LLC | 臨床1/2期 |
| TQB-2223/Penpulimab | 單克隆抗體 | 晚期癌癥 | 正大天晴藥業(yè)集團股份有限公司 | 臨床1期 |
| AK137 | 雙特異性抗體 | 晚期癌癥 | 中山康方生物醫(yī)藥有限公司 | 臨床1期 |
| INCA32459 | 雙特異性抗體 | 轉(zhuǎn)移性頭頸部鱗狀細(xì)胞癌 | 不可切除的黑色素瘤 | Incyte Corp. | 臨床1期 |
| IBI-323 | 雙特異性抗體 | 晚期癌癥 | 信達(dá)生物制藥(蘇州)有限公司 | 臨床1期 |
| IMP-761 | 單克隆抗體 | 自身免疫性疾病 | Immutep Ltd. | 臨床1期 |
| ABL-501 | 雙特異性抗體 | 晚期惡性實體瘤 | ABL Bio, Inc. | 臨床1期 |
| Negalstobart | 單克隆抗體 | 彌漫性大B細(xì)胞淋巴瘤 | 晚期癌癥 | 胃食管交界處癌 | 鱗狀非小細(xì)胞肺癌 | 鼻咽癌 | 信達(dá)生物制藥(蘇州)有限公司 | 臨床1期 |
| TQB-2223 | 單克隆抗體 | 晚期肝細(xì)胞癌 | 晚期癌癥 | 膽管癌 | Vater壺腹癌 | 食管鱗狀細(xì)胞癌 | 膽囊癌 | 復(fù)發(fā)性實體腫瘤 | 腫瘤轉(zhuǎn)移 | Symphogen A/S | 明日陽光藥業(yè)公司 | 正大天晴藥業(yè)集團股份有限公司 | Clarusun Pharmaceutical Corp. | 臨床1期 |
| ZGGS15 | 雙特異性抗體 | 晚期惡性實體瘤 | 實體瘤 | 蘇州澤璟生物制藥股份有限公司 | 臨床1期 |
| 重組人源化單克隆抗體MIL98 | 單克隆抗體 | 晚期淋巴瘤 | 黑色素瘤 | 北京天廣實生物技術(shù)股份有限公司 | 臨床1期 |
| BJ-007 | 單克隆抗體 | 感染 | 上海寶濟藥業(yè)股份有限公司 | 臨床申請批準(zhǔn) |
(數(shù)據(jù)來源于patsnap)
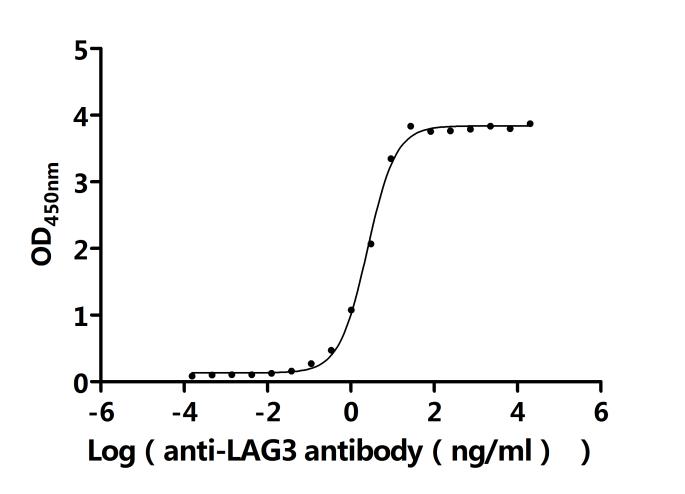
6. 華美生物L(fēng)AG3相關(guān)產(chǎn)品推薦
LAG3作為關(guān)鍵免疫檢查點,通過與TCR-CD3復(fù)合物結(jié)合、干擾共受體信號及與配體相互作用,在腫瘤免疫逃逸、自身免疫疾病及慢性感染中發(fā)揮重要作用。華美生物提供LAG3 重組蛋白、抗體、ELISA試劑盒產(chǎn)品,助力您的機制研究或藥物開發(fā)。

參考文獻(xiàn):
[1] Nguyen L T, Ohashi P S. Clinical blockade of PD1 and LAG3--potential mechanisms of action[J]. Nat Rev Immunol, 2015, 15(1): 45-56.
[2] Hua S, et al. LAG3 (CD223) and autoimmunity: Emerging evidence[J]. J Autoimmun, 2020, 114: 102504.
[3] Guy C, et al. LAG3 associates with TCR-CD3 complexes and suppresses signaling by driving co-receptor-Lck dissociation[J]. Nat Immunol, 2022, 23(5): 757-767.
[4] Ming Q, et al. LAG3 ectodomain structure reveals functional interfaces for ligand and antibody recognition[J]. Nat Immunol, 2022, 23(7): 1031-1041.
[5] Singh L, et al. Prognostic significance of lymphocyte activation gene-3 (LAG3 gene) in uveal melanoma patients[J]. Annals of Oncology, 2021, 32(S7): S1463.
[6] Gurler F, et al. Retrospective analysis of PDL-1, LAG3, TIM3, OX40L and MSI status in gastroenteropancreatic neuroendocrine tumors (GEP-NETs)[J]. Annals of Oncology, 2021, 32(S5): S622.
[7] Burugu S, et al. LAG-3+ tumor infiltrating lymphocytes in breast cancer: clinical correlates and association with PD-1/PD-L1+ tumors[J]. Ann Oncol, 2017.
[8] LAG3 landscape in solid tumors and its association with immunotherapy outcomes in non-small cell lung cancer.
[9] LAG3-related factors to predict response to nivolumab monotherapy in advanced gastric cancer (WJOG10417GTR study).
[10] Phase 1 trial of the anti-LAG3 antibody favezelimab plus pembrolizumab in advanced gastric cancer.
[11] LBA31 Neoadjuvant nivolumab plus relatlimab (anti-LAG3) in locally advanced MMR-deficient colon cancers The NICHE-3 study.
[12] Xiu-Qing P, et al. LAG3+ erythroid progenitor cells inhibit HBsAg seroclearance during finite pegylated interferon treatment through LAG3 and TGF-β[J]. Antiviral Res, 2023.









