蛋白質乙酰化: 真核細胞中的一種重要翻譯后修飾
日期:2024-02-02 16:03:52
眾所周知,翻譯后修飾(PTM)是真核細胞實現蛋白質功能多樣化和動態協調信號網絡的重要機制。1964年,Vincent Allfrey及其同事首次發現了組蛋白上的賴氨酸乙酰化修飾。隨后,以質譜儀為基礎的蛋白質組學大大加速了內源性乙酰化蛋白質的發現和鑒定,同時也揭示了非組蛋白乙酰化的調控過程。乙酰化修飾是一種進化保守的PTM,存在于原核生物和真核生物中。下文將重點介紹蛋白質乙酰化的定義、功能、機制以及相關疾病。
1. 什么是蛋白質乙酰化?
蛋白質乙酰化是真核生物中主要的翻譯后修飾(PTMs)之一,其中乙酰輔酶A(Ac-CoA)的乙酰基被引入多肽鏈上的特定位點 [1]。蛋白質被乙酰化的部位要么是各種氨基末端殘基,要么是賴氨酸殘基的ε-氨基。大多數真核生物蛋白質和調節肽在氨基末端殘基上發生乙酰化,而賴氨酸乙酰化則發生在組蛋白和轉錄因子等不同蛋白質的不同位點上。
2. 蛋白質乙酰化的功能
人類的大多數蛋白質都會發生乙酰化。蛋白質的乙酰化可影響蛋白質的電荷、構象、穩定性、定位、合成以及與其他分子的相互作用。已發現許多乙酰化蛋白質參與了多種細胞過程,如翻譯、轉錄、蛋白質折疊、細胞分裂、DNA損傷修復、信號轉導和新陳代謝。組蛋白的乙酰化可降低其正電荷,削弱其與DNA結合的能力,并導致核小體解聚,從而使轉錄因子和RNA聚合酶順利與DNA結合,激活基因的轉錄活性。研究發現,乙酰化可影響核酸酶的酶活性,從而調節底物RNA的水平 [7]。乙酰化還能調節100多種非組蛋白,包括轉錄因子、轉錄輔激活因子和核受體 [8]。非組蛋白乙酰化使細胞功能復雜化,而線粒體關鍵酶的乙酰化則調節生物能代謝。蛋白質乙酰化還與蛋白質降解有關。此外,蛋白質乙酰化還能調節多種信號通路并影響細胞周期。
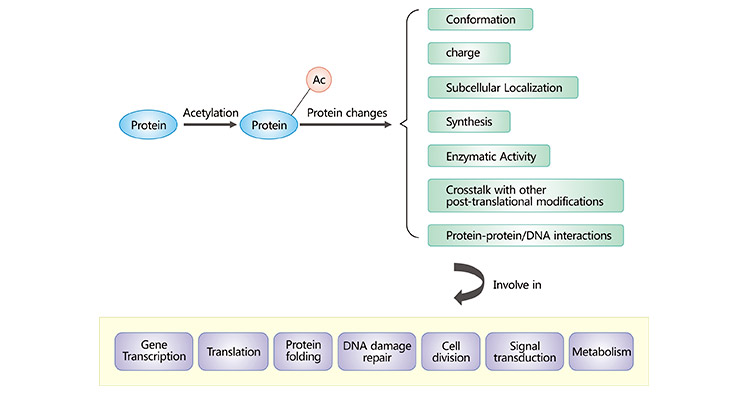
圖1. 蛋白質乙酰化在真核細胞中的作用
3. 蛋白質乙酰化的機制
研究最多的蛋白質乙酰化發生在氨基上,但也檢測到絲氨酸、蘇氨酸和組氨酸殘基上的乙酰化。蛋白質氨基的乙酰化有三種不同的機制:賴氨酸乙酰化(N?-乙酰化)、蛋白質N端乙酰化(N α-乙酰化)和O-乙酰化。
3.1 N?-賴氨酸乙酰化
賴氨酸乙酰化又稱 N?-賴氨酸乙酰化,是指將乙酰基從乙酰輔酶 A(乙酰輔酶 A)轉移到蛋白質中賴氨酸側鏈 ? 位的伯胺上。這一可逆過程中和了賴氨酸側鏈?位的正電荷。失去正電荷和增大的賴氨酸會破壞鹽橋并引入立體結構,從而改變蛋白質-蛋白質/DNA 之間的相互作用、穩定性和酶活性 [2][3]。
有力的證據表明,蛋白質賴氨酸乙酰化可通過兩種不同的機制發生:酶促乙酰化和非酶促乙酰化(化學乙酰化)[4]。兩種不同的機制顯示了對不同賴氨酸乙酰化位點的偏好,并揭示了這些賴氨酸位點相對乙酰化變化的不同動態。
酶促乙酰化依靠乙酰輔酶 A(Ac-CoA)乙酰轉移酶催化乙酰基向賴氨酸殘基的?-氨基轉移。非酶促乙酰化發生在乙酰供體和蛋白質之間。在真核細胞中,特別是在線粒體內,高能硫代酯類 AcCoA 可對蛋白質進行化學乙酰化 [16]。
3.2 Nα-乙酰化
Nα-乙酰化是指在N端氨基酸的 α-氨基上添加乙酰基。這是一個典型的不可逆過程,由 N-α-乙酰轉移酶(NATs)介導。大約85%的人類蛋白質是通過 Nα-乙酰化修飾的 [5]。在真核生物中,Nα-乙酰化非常常見,而且是翻譯過程中的共同作用,但在細菌中卻很少見,而且是翻譯后的作用 [17]。在真核細胞中,Nα-乙酰化要么發生在蛋氨酸的游離氨基上,要么發生在N端蛋氨酸裂解后暴露的氨基酸上。在細菌、線粒體和葉綠體中,Nα-乙酰化是翻譯后進行的,因為蛋氨酸必須首先進行變形。
3.3 O-乙酰化
O-乙酰化是指在酪氨酸/絲氨酸/蘇氨酸殘基的羥基上添加乙酰基 [6]。它已被確定為蛋白質乙酰化的第三種類型。耶爾森氏菌外層蛋白J(YopJ)會對激活MAPK/ERK激酶和IκB激酶家族所需的絲氨酸和蘇氨酸殘基側鏈進行乙酰化,從而阻斷它們的磷酸化,抑制信號轉導。蛋白質N-聚糖中的硅醛酸的O-乙酰化是一種重要的修飾,可以在 4-、7-、8-或9-位上以不同的組合出現。
4. 蛋白質乙酰化與疾病
功能蛋白的乙酰化和去乙酰化在胚胎發育、出生后成熟、心肌細胞分化、心臟重塑以及各種心血管疾病(包括肥胖、糖尿病、心臟代謝性疾病、缺血再灌注損傷以及心臟重塑、高血壓和心律失常等)的發病過程中發揮著重要作用 [10] [11]。
歐陽潔等研究表明,腎小管上皮細胞線粒體中 SOD2 和 p53 蛋白乙酰化的升高是缺血再灌注(I/R)誘導急性腎損傷(AKI)發病機制中的一個重要信號事件 [9]。一些研究還表明,恢復 SIRT1/3 的活性可能是治療 AKI 的新靶點。SIRT1 和 3 是依賴于煙酰胺腺嘌呤二核苷酸的蛋白去乙酰化酶,在抗氧化應激和抗細胞凋亡過程中發揮保護腎功能的作用。使用白藜蘆醇可以有效恢復SIRT1/3的活性。研究表明,TDP-43的K136乙酰化會損害其RNA結合和剪接能力,促進病理磷酸化和泛素化TDP-43的不溶性聚集體的積累,這與肌萎縮性脊髓側索硬化癥(ALS)有關。
某些蛋白質的乙酰化與癌變有關。與相應的非腫瘤組織細胞相比,結腸癌相關轉錄因子1(CCAT1)在食管鱗狀細胞癌(ESCC)細胞中的表達明顯更高 [12]。眾所周知,CCAT1的高表達可促進細胞增殖和侵襲,而下調 CCAT1 則可抑制這兩個生物學過程 [13]。據報道,H3K27的乙酰化可部分上調CCAT1的表達,而CCAT1有可能誘發癌癥 [14]。此外,糖酵解為癌細胞快速增殖提供了大量能量。磷酸甘油酸激酶1(PGK1)是糖酵解過程中的重要還原酶,如果發生乙酰化,可能會改變癌細胞的增殖。在肝癌細胞中,PGK1的活性會因乙酰化而增強,并進一步加速腫瘤細胞的增殖 [15]。
質譜技術與生物學的結合使大量乙酰化位點在所有蛋白質中的定位成為可能。越來越多的研究表明,乙酰化是自然界中最豐富的化學修飾之一,可能影響蛋白質的各種生理過程,甚至導致某些疾病。因此,蛋白質乙酰化也是近年來開發和設計治療多種疾病的新藥的一個很有前景的靶點。
5. 乙酰化相關產物
許多乙酰轉移酶都參與了蛋白質的乙酰化過程。在此,CUSABIO為科研人員提供一些優質的乙酰轉移酶,用于蛋白質乙酰化相關的研究。
| 乙酰化蛋白 | 乙酰轉移酶 |
|---|---|
| Human proteins | NatA (catalytic subunit: Naa10, Naa15 ), NatB (Naa20, Naa25), NatC (Naa30, Naa35, Naa38), NatD (Naa40), NatE (Naa50), NatF (Naa60), NatH (Naa80) |
| Histones (H2A, H2B, H3, H4) | Gcn5, PCAF, Hat1, Elp3, Hpa2, Esa1, MOF, Sas2, Sas3, Tip60, MORF, TAFII250, TFIIIC, ACTR, and SRC1. |
| E. coli ribosomal proteins (S18, S5, and L12) | RimI, RimJ, RimL |
| Transcription factors (p53, E2F1-3, EKLF) | PCAF/Gcn5, p300/CBP, TAFII250 |
| Nuclear import factors (importin-α7 and Rch1) | p300/CBP |
參考文獻:
[1] Adrian Drazica, Line M.Myklebust, et al. The world of protein acetylation [J]. Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics Volume 1864, Issue 10, October 2016, Pages 1372-1401.
[2] David G. Christensen, Xueshu Xie, et al. Post-translational Protein Acetylation: An Elegant Mechanism for Bacteria to Dynamically Regulate Metabolic Functions [J]. Front Microbiol. 2019; 10: 1604.
[3] Ibraheem Ali, Ryan J. Conrad, et al. Lysine Acetylation Goes Global: From Epigenetics to Metabolism and Therapeutics [J]. Chem Rev. Author manuscript; available in PMC 2019 Jul 3.
[4] Miao-Miao Wang, Di You & Bang-Ce Ye. Site-specific and kinetic characterization of enzymatic and nonenzymatic protein acetylation in bacteria [J]. Sci Rep 7, 14790 (2017).
[5] Hollebeke J, Van Damme P and Gevaert K: N-terminal acetylation and other functions of Nalpha-acetyltransferases [J]. Biol Chem. 393:291–298. 2012.
[6] Yang XJ and Gregoire S. Metabolism, cytoskeleton and cellular signalling in the grip of protein Nepsilon- and O-acetylation [J]. EMBO Rep. 8:556–562. 2007.
[7] Song L, Wang G, et al. Reversible acetylation on Lys501 regulates the activity of RNase II [J]. Nucleic Acids Res. 2016;44:1979–1988.
[8] Narita T, Weinert BT, et al. Functions and mechanisms of non-histone protein acetylation [J]. Nat Rev Mol Cell Biol. 2019;20:156–174.
[9] Jie Ouyang, Zhenhua Zeng, et al. SIRT3 Inactivation Promotes Acute Kidney Injury Through Elevated Acetylation of SOD2 and p53 [J]. J Surg Res. 2019 Jan;233:221-230.
[10] Can Xia, Yu Tao, et al. Protein acetylation and deacetylation: An important regulatory modification in gene transcription (Review) [J]. Exp Ther Med. 2020 Oct; 20(4): 2923–2940.
[11] MingjieYang, YingmeiZhang, et al. Acetylation in cardiovascular diseases: Molecular mechanisms and clinical implications [J]. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease Volume 1866, Issue 10, 1 October 2020, 165836.
[12] Hu M, Zhang Q, et al. lncRNA CCAT1 is a biomarker for the proliferation and drug resistance of esophageal cancer via the miR- 143/PLK1/BUBR1 axis [J]. Mol Carcinog. 2019;58:2207–2217.
[13] Li J and Qi Y. Ginsenoside Rg3 inhibits cell growth, migration and invasion in Caco-2 cells by downregulation of lncRNA CCAT1 [J]. Exp Mol Pathol. 2019;106:131–138.
[14] Zhang E, Han L, et al. H3K27 acetylation activated-long non-coding RNA CCAT1 affects cell proliferation and migration by regulating SPRY4 and HOXB13 expression in esophageal squamous cell carcinoma [J]. Nucleic Acids Res. 2017;45:3086–3101.
[15] Hu H, Zhu W, et al. Acetylation of PGK1 promotes liver cancer cell proliferation and tumorigenesis [J]. Hepatology. 2017;65:515–528.
[16] Hosp F, Lassowskat I, et al. Lysine acetylation in mitochondria: from inventory to function [J]. Mitochondrion 2017. 33:58–71.
[17] Soppa J. Protein acetylation in archaea, bacteria, and eukaryotes [J]. Archaea 2010:820681.
上一篇: 蛋白質磷酸化









