Recombinant Rat Vesicular glutamate transporter 1 (Slc17a7), partial
-
中文名稱:大鼠Slc17a7重組蛋白
-
貨號:CSB-EP730797RA-B
-
說明書:
-
規格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
產品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
別名:Slc17a7; Bnpi; Vglut1Vesicular glutamate transporter 1; VGluT1; Brain-specific Na(+)-dependent inorganic phosphate cotransporter; Solute carrier family 17 member 7
-
種屬:Rattus norvegicus (Rat)
-
蛋白長度:Partial
-
蛋白標簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產品提供形式:Lyophilized powder Warning: in_array() expects parameter 2 to be array, null given in /www/web/cusabio_cn/public_html/caches/caches_template/default/content/show_product_protein.php on line 662
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產品評價

相關產品
靶點詳情
-
功能:Mediates the uptake of glutamate into synaptic vesicles at presynaptic nerve terminals of excitatory neural cells. May also mediate the transport of inorganic phosphate.
-
基因功能參考文獻:
- findings indicate that unilateral cochlear ablation suppresses VGLUT1 expression in the central auditory pathways not only ipsilateral but also contralateral to the ablated side. PMID: 28238468
- VGLUT1 was transiently down-regulated in ipsilateral dorsal root ganglia and contralateral spinal cord in a model of spared nerve injury. PMID: 27210824
- In previously unstressed rats, vGluT1 siRNA significantly enhanced ACTH and corticosterone secretion. Compared with chronic variable stress (CVS) animals receiving the green fluorescent protein control vector, the vGluT1 siRNA further increased basal and stress-induced corticosterone release. Further analysis revealed enhanced adrenal responsiveness in CVS rats treated with vGluT1 siRNA. PMID: 28938481
- results indicated that VGLUT1v exists in a functional state in rat photosensitive cells and is involved in glutamatergic chemical transmission PMID: 28188742
- vGlut1 has a strong influence on the kinetics of retrieval of half of the known SV cargos and that specifically impairing the endocytosis PMID: 26224632
- The results indicated that glutamatergic spinal trigeminal nucleus neurons sending their axons to the cerebellar cortical regions mainly express VGLUT1. PMID: 23380804
- Following peripheral nerve injury, 62% of VGLUT1 synapses are lost on spinal motor neuron dendrites. PMID: 24599449
- VGLUT1 contains multiple sorting signals that engage distinct trafficking mechanisms PMID: 23804088
- These results suggest that BDNF regulates VGLUT expression during development and its effect on VGLUT1 may contribute to enhance glutamate release in LTP PMID: 23326507
- Up-regulation of VGLUT1 is important for development of presynaptic terminals in the cortex. PMID: 23226425
- VGluT1 immunoreactions are localized within the cytoplasm for all sizes of trigeminal neurons, although predominantly in medium-large trigeminal neurons. PMID: 21957077
- VGLUT1 labeling is concentrated in astrocytic processes from wild type, and not in VGLUT1 knockout hippocampus. PMID: 22009457
- Expression of VGLUT2 precedes that of VGLUT1 during olfactory bulb morphogenesis, suggesting differential contribution of the two VGLUT subtypes to glutamate-mediated embryonic events. PMID: 21609737
- After nerve injuries synapses are stripped from motoneurons, other excitatory and inhibitory inputs eventually recover but VGLUT1 synapses are permanently lost on the cell body (75-95% synaptic losses) and on the proximal 100 mum of dendrite (50% loss). PMID: 21832035
- The VGLUT1 upstream promoter and the first intron, fused to the basal promoter, both support glutamatergic-specific expression in rat postrhinal cortex. PMID: 21172319
- The vesicular glutamate transporter-1 (VGLUT1)promoter supports expression in VGLUT1-containing glutamatergic neurons in rat postrhinal cortex. PMID: 20849834
- Most of the glutamatergic neurons that expressed glutamate vesicular transporter 1 also co-expressed choline acetyltransferase. PMID: 20632124
- There are regionally significant correlations between VGLUT1 concentration and cognitive scores between different dementia groups and those with stroke who do not develop dementia. PMID: 21079182
- observed a biphasic and bilateral change in the protein expression levels of both VGLUTs in the striatum of hemi-Parkinson rats indicative for a different and time-dependent change in glutamatergic neurotransmission from two types of striatal afferents PMID: 20450947
- The study suggested that VGLUT enhances transmitter storage in nonglutamatergic neurons. PMID: 20519538
- The presence of the glutamate markers VGLUT1 and VGLUT2 in distinct populations of peptide hormone-secreting hypophysial cells highly indicates the involvement of endogenous glutamate release in autocrine/paracrine regulatory mechanisms PMID: 20025917
- These findings show the specific lingual pattern of distribution for VGluts and suggest that only VGlut 1 takes part in glutamatergic regulation of epithelial and taste cells within the tongue in physiological conditions. PMID: 19952853
- VGLUT1 immunoreactivity was detected in the sensory endings on the equatorial and juxta-equatarial regions of intrafusal fibers as well as in many axon terminals within lamina IX of the spinal cord. PMID: 15157812
- VGLUT1 provides a specific marker for glutamatergic neuronal elements in the septal hippocampus. PMID: 15224985
- The developmental pattern of Vglut1 in the olivary nucleus was studied. PMID: 15714284
- the coupling of zinc transporter 3 and Vglut1 transport mechanisms regulates neurotransmitter content in secretory vesicles PMID: 15860731
- A newly proposed mechanism for altering the strength of glutamatergic synapses based on the recently identified vesicular glutamate transporter VGLUT1. PMID: 15987952
- a high proportion of cardiac neurones appear to be glutamatergic, but differ from other vagal afferents in expressing vesicular glutamate transporter 1 PMID: 16084661
- VGLUT1-ir was found in extrinsic as well as intrinsic innervation of the rat esophagus. PMID: 16231188
- Overall, the discovery of endophilin as a partner for VGLUT1 in nerve terminals strongly suggests the existence of functional differences between VGLUT1 and -2 terminals in their abilities to replenish vesicle pools. PMID: 16606361
- VGLUT1 and endophilins may interact to play an important role affecting the vesicular sequestration and synaptic release of glutamate. PMID: 16710756
- Lamina I and IIo displayed a moderate density of small VGLUT1 varicosities at all spinal levels. Corticospinal tract axons displayed VGLUT1, indicating that the corticospinal tract is an important source of small VGLUT1 varicosities. PMID: 16786558
- After stimulation, inhibition of the AP3 pathway prevents the full recovery of VGLUT1 by endocytosis, implicating the AP3 pathway specifically in compensatory endocytosis. PMID: 16815333
- These data indicate that VGLUT1 is expressed in both the central axon terminals and the peripheral sensory endings of Vmes neurons, although no VGLUT1 immunoreactivity was detectable in the cell bodies of Vmes neurons in adult rats. PMID: 16856164
- In situ hybridization with variant-specific probes showed expression of VGLUT1v in the inner segment layer of photoreceptor cells. PMID: 16987242
- No significant change in either VGLUT1 or GlyT2 labeling of sacrocaudal motoneurons at any of the time points examined. PMID: 17134699
- There was a striking loss of mossy cells during the latent period, demonstrated by a long-term decrease of VGLUT1 mRNA-containing hilar neurons and associated loss of VGLUT1-containing terminals in the dentate gyrus. PMID: 17503488
- VGluT1 was localized to partial neurons of VG and to the putative primary afferent fibers innervating vestibular end-organs. PMID: 17612597
- Histological analysis revealed that intensities of VGLUT1 hybridization signal and immunostaining drastically increase by postnatal day (P) 7 in pinal gland. PMID: 18291592
- Results showed numerous VGLUT1 immunolabeled terminals in the central nucleus, lateral cortex and dorsal cortex, and Results showed numerous VGLUT1 and 2 immunolabeled terminals in the central nucleus, lateral cortex and dorsal cortex. PMID: 18436385
- In rat retina, VGLUT1 was initially detected at postnatal day (P5) in photoreceptor terminals and P6 in bipolar terminals. PMID: 18482716
- In the present study observed a higher expression of vglut1 mRNA in the left cingulate cortex. This suggests a hemispheric asymmetry in the strength of the cortico-subcortical glutamatergic system. PMID: 19626270
- In cortical cultures and layer V cortical terminals VGLUT1 is coexpressed with vesicular transporter VGAT in the same population of synaptic vesicles. PMID: 19627441
顯示更多
收起更多
-
亞細胞定位:Cytoplasmic vesicle, secretory vesicle, synaptic vesicle membrane. Membrane; Multi-pass membrane protein. Cell junction, synapse, synaptosome.
-
蛋白家族:Major facilitator superfamily, Sodium/anion cotransporter family, VGLUT subfamily
-
組織特異性:Expressed in the cerebellum, cerberal cortex and hippocampus. Isoform 2 is expressed specifically in retina.
-
數據庫鏈接:
Most popular with customers
-
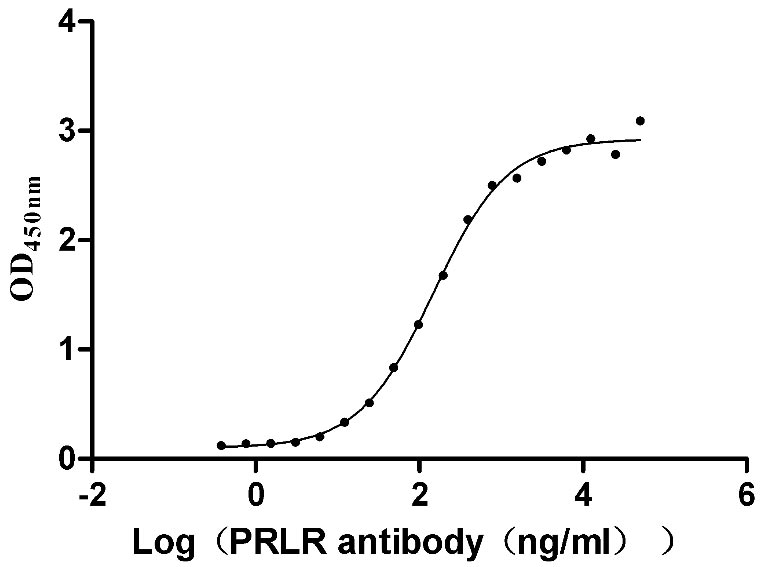
Recombinant Human Prolactin receptor (PRLR), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
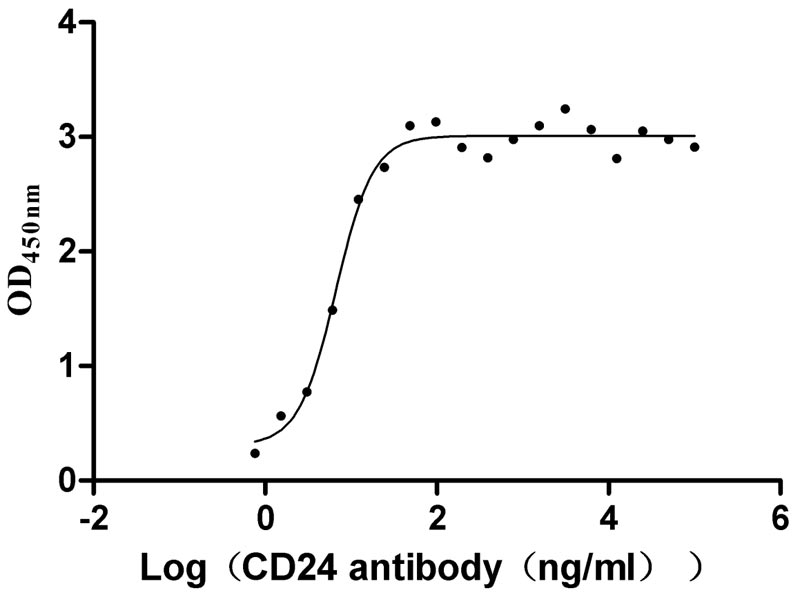
Recombinant Human Signal transducer CD24 (CD24)-Nanoparticle (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
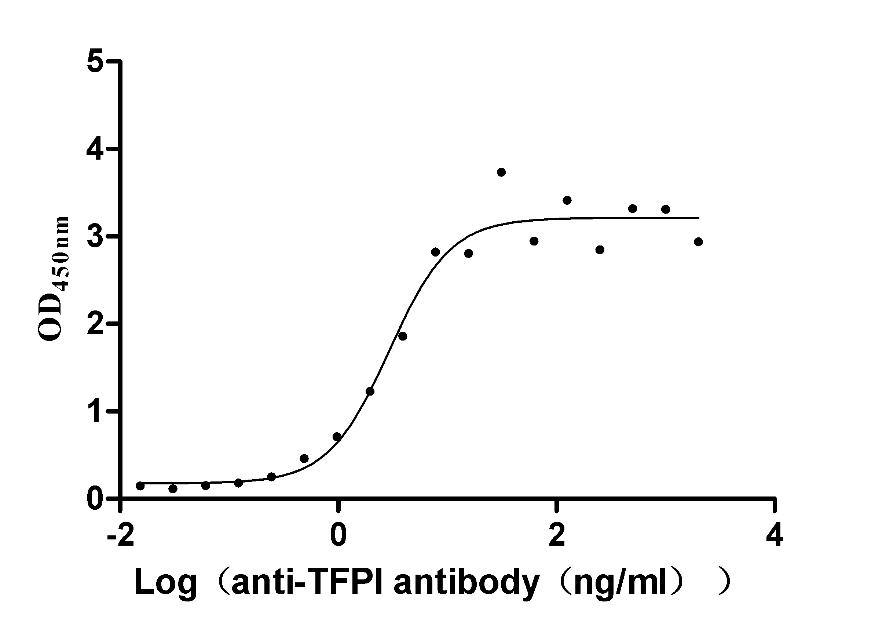
Recombinant Rabbit Tissue factor pathway inhibitor (TFPI) (Active)
Express system: Mammalian cell
Species: Oryctolagus cuniculus (Rabbit)
-
f4-AC1.jpg)
Recombinant Human Claudin-6 (CLDN6)-VLPs, Fluorescent (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Urokinase-type plasminogen activator(PLAU) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Human Mucin-13(MUC13),partial (Active)
Express system: yeast
Species: Homo sapiens (Human)
-
Recombinant Human Tyrosine-protein kinase receptor UFO(AXL),partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human C-C chemokine receptor type 5 (CCR5)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)









