Recombinant Mouse Granulocyte-macrophage colony-stimulating factor (Csf2)
-
中文名稱:Recombinant Mouse Granulocyte-macrophage colony-stimulating factor(Csf2)
-
貨號(hào):CSB-YP006045MO
-
說(shuō)明書(shū):
-
規(guī)格:
-
來(lái)源:Yeast
-
其他:
-
中文名稱:Recombinant Mouse Granulocyte-macrophage colony-stimulating factor(Csf2)
-
貨號(hào):CSB-EP006045MO-B
-
說(shuō)明書(shū):
-
規(guī)格:
-
來(lái)源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:Recombinant Mouse Granulocyte-macrophage colony-stimulating factor(Csf2)
-
貨號(hào):CSB-BP006045MO
-
說(shuō)明書(shū):
-
規(guī)格:
-
來(lái)源:Baculovirus
-
其他:
產(chǎn)品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
別名:Csf2; Csfgm; Granulocyte-macrophage colony-stimulating factor; GM-CSF; Colony-stimulating factor; CSF
-
種屬:Mus musculus (Mouse)
-
蛋白長(zhǎng)度:Full Length of Mature Protein
-
表達(dá)區(qū)域:18-141
-
氨基酸序列APT RSPITVTRPW KHVEAIKEAL NLLDDMPVTL NEEVEVVSNE FSFKKLTCVQ TRLKIFEQGL RGNFTKLKGA LNMTASYYQT YCPPTPETDC ETQVTTYADF IDSLKTFLTD IPFECKKPGQ K
-
蛋白標(biāo)簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產(chǎn)品提供形式:Lyophilized powder Warning: in_array() expects parameter 2 to be array, null given in /www/web/cusabio_cn/public_html/caches/caches_template/default/content/show_product_protein.php on line 662
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復(fù)溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲(chǔ)存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質(zhì)期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項(xiàng):Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產(chǎn)品評(píng)價(jià)

相關(guān)產(chǎn)品
靶點(diǎn)詳情
-
功能:Cytokine that stimulates the growth and differentiation of hematopoietic precursor cells from various lineages, including granulocytes, macrophages, eosinophils and erythrocytes.
-
基因功能參考文獻(xiàn):
- Data show that the microbiota enhances respiratory defenses via granulocyte-macrophage colony-stimulating factor (GM-CSF) signaling, which stimulates pathogen killing and clearance by alveolar macrophages PMID: 29142211
- The data indicate that GM-CSF drives chronic tissue damage and disability in experimental autoimmune encephalomyelitis via pleiotropic pathways, but it is dispensable during early lesion formation and the onset of neurologic deficits. PMID: 29288202
- the release of IL-33 and GM-CSF from epithelial cells induces the activation of p65 and the p38-MK2/3 signaling module in Dendritic Cells, resulting in Th2 polarization and, finally, allergic inflammation. PMID: 29288203
- results show T cell production of GM-CSF contributes to control of M. tuberculosis infection in the absence of other sources of GM-CSF, that multiple T cell subsets make GM-CSF in the lung over the course of infection and that GM-CSF can act directly on infected macrophages through a pathway requiring PPARgamma to limit bacterial growth PMID: 29066547
- In conclusion, our study confirms the pathogenic role of GM-CSF in colitis-associated colorectal cancer development. GM-CSF favors tumor-permissive microenvironment by inducing MDSC generation and recruiting them into colonic tissues. PMID: 28534709
- these data demonstrate that GM-CSF levels during radiotherapy can be used as a prognostic biomarker for lung and esophageal cancer PMID: 27835886
- this study demonstrates that epithelial-derived GM-CSF is a critical early signal during allergic sensitization to cockroach allergen PMID: 27731325
- These impaired macrophage functions in leukemic mice were significantly corrected by IL-3 and GM-CSF treatment indicating the therapeutic benefit of these two cytokines in leukemia. PMID: 28039843
- Both IL-6 protein production and transcript levels were downregulated by RA in respiratory tract epithelial cells (LETs) , but upregulated in macrophages (MACs). RA also increased transcript levels of MCP-1, GMCSF, and IL-10 in MACs, but not in LETs. Conversely, when LETs, but not MACs, were exposed to RA PMID: 27940088
- T-GM-CSF and -IL-3 significantly, and reciprocally, blunted receptor binding and myeloid progenitor cell proliferation activity of both FL-GM-CSF and -IL-3 in vitro and in vivo PMID: 28344320
- Results indicate GM-CSF as both a key contributor to the pathogenesis of MI and a potential therapeutic target. PMID: 28978634
- Obesity alters the lung neutrophil infiltration to enhance breast cancer metastasis through IL5 and GM-CSF. PMID: 28737771
- GM-CSF is required for the normal balance of leukocyte subsets, including granulocytes, B cells, and naive vs. effector T cells. There was an approximately 3-fold increase in the percentages of granulocytes in Csf2-/- PBMCs. The presence of maximal experimental autoimmune encephalomyelitis in the complete absence of GM-CSF revealed that GM-CSF is not an obligate effector molecule in all forms of EAE. PMID: 27256565
- chemerin inhibited nuclear factor-kappaB activation and the expression of granulocyte-macrophage colony-stimulating factor (GM-CSF) and interleukin-2 (IL-6) by tumor cells and tumor-associated endothelial cell, respectively, via its receptors, and consequently, MDSC induction was impaired, leading to restoration of antitumor T-cell response and decreased tumor angiogenesis. PMID: 28166197
- These findings describe a novel role for GM-CSF as an essential initiating cytokine in cardiac inflammation. PMID: 27595596
- Data reviewed establish that any damage to brain tissue tends to cause an increase in G-CSF and/or GM-CSF (G(M)-CSF) synthesized by the brain. Glioblastoma cells themselves also synthesize G(M)-CSF. G(M)-CSF synthesized by brain due to damage by a growing tumor and by the tumor itself stimulates bone marrow to shift hematopoiesis toward granulocytic lineages away from lymphocytic lineages. PMID: 28459367
- Evi1(+)DA-3 cells modified to express an intracellular form of GM-CSF, acquired growth factor independence and transplantability and caused an overt leukemia in syngeneic hosts, without increasing serum GM-CSF levels. PMID: 25907616
- IL-23-induced GM-CSF mediates the pathogenicity of CD4(+) T cells in experimental autoimmune myocarditis. PMID: 26660726
- GM-CSF accelerated the G1/S phase transition in EPCs by upregulating the expression of cyclins D1 and E. PMID: 24662605
- host RNF13 affects the concentration of GM-CSF in tumor-bearing lungs PMID: 26197965
- Sc CW-derived BG stimulated the late and strong expression of Csf2 in a dectin-1-dependent manner, they remain poor inducers of chemokine and cytokine production in murine macrophages. PMID: 26840954
- GM-CSF and uPA are required for Porphyromonas gingivalis-induced alveolar bone loss in a mouse periodontitis model. PMID: 25753270
- These findings identify GM-CSF as central to the protective immune response that prevents progressive fungal disease PMID: 26755822
- regulatory molecule causative of aortic dissection and intramural haematoma PMID: 25923510
- Results indicate that granulocyte-macrophage colony-stimulating factor (GM-CSF) signaling controls a pathogenic expression signature in CC chemokine receptors 2 (CCR2+)Ly6C(hi) monocytes. PMID: 26341401
- TL1A increases expression of CD25, LFA-1, CD134 and CD154, and induces IL-22 and GM-CSF production from effector CD4 T-cells PMID: 25148371
- Mafb-deficient microglia cultured with GM-CSF barely extended their membrane protrusions, probably due to abnormal activation of RhoA, a key regulator of cytoskeletal remodeling. MafB is a negative regulator of GM-CSF signaling in microglia. PMID: 25998393
- Csf2-/- mice showed a 30% increase in whole body adiposity, which persisted to adulthood. Adipocytes from Csf2-/- mice were 50% larger by volume and contained higher levels of adipogenesis gene transcripts, indicating enhanced adipocyte differentiation. PMID: 25931125
- Renal ischemia reperfusion injury tubular cells expressed elevated GM-0CSF, that supports tubular cells proliferation. PMID: 25388222
- GM-CSF is key to the development of experimental osteoarthritis and its associated pain. PMID: 22995428
- Loss of encephalitogenic activity of STAT5-deficient autoreactive CD4+ T cells was independent of IFN-gamma or IL-17 production, but was due to the impaired expression of GM-CSF, a crucial mediator of T-cell pathogenicity. PMID: 25412660
- These data indicate that GM-CSF plays a role in the inflammatory signaling network that drives neutrophil recruitment in response to Clostridium difficile infection but does not appear to play a role in clearance of the infection. PMID: 25045999
- IRF8 represses GM-CSF expression in T cells to affect myeloid cell lineage differentiation. PMID: 25646302
- Reprogramming of monocytes by GM-CSF contributes to regulatory immune functions during intestinal inflammation. PMID: 25653427
- GM-CSF promotes advanced plaque progression by increasing macrophage apoptosis susceptibility. PMID: 25348165
- although iTreg cells maintained the ability to produce IFN-gamma and TNF-alpha in vivo, their ability to produce GM-CSF was selectively degraded upon antigen stimulation under inflammatory conditions. PMID: 25168419
- IFN-gamma has a pprotective role in the demyelination of brain through downregulation of IL-17/GM-CSF and induction of neuroprotective factors in the brain by monocytes/microglial cells. PMID: 25339670
- Expression of PPAR-gamma in fetal lung monocytes was dependent on the cytokine GM-CSF. Therefore, GM-CSF has a lung-specific role in the perinatal development of alveolar macrophages through the induction of PPAR-gamma in fetal monocytes. PMID: 25263125
- These novel findings indicate that the inflammatory mediator, GM-CSF, may have significant protective properties in the chronic sequelae of experimental Traumatic brain injury PMID: 24392832
- Airway infections and pollutants increase the susceptibility to develop allergic asthma through a GM-CSF/IL-33/OX40L pathway. PMID: 24551140
- Data indicate that neutralization of GM-CSF would be a useful therapeutic strategy for severe connective tissue diseases (CTD)-interstitial lung disease (ILD). PMID: 24951817
- Data indicate that lung epithelium-derived GM-CSF is a critical regulator of CD11b+ dendritic cells (DCs)-mediated Th2 cell priming. PMID: 24943219
- Bhlhe40 is required positively regulates the production of GM-CSF and negatively regulates the production of IL-10 in T cells. PMID: 24699451
- Report IL-18 plus GM-CSF modified tumor cell vaccine can induce significant antitumor effects in Lewis lung carcinoma. PMID: 24475975
- Data indicate that in type 1 diabetes monocytes STAT5Ptyr activation is significantly higher and is found bound to CSF2 promoter and PTGS2 enhancer regions. PMID: 24204704
- Data indicate that GM-CSF controls IgM production in innate response activator B cells. PMID: 24821911
- our findings suggest that over-production of GM-CSF by T cells could be a pathogenic factor in many histiocytic disorders PMID: 24183847
- that p52 binds to the promoter of the GM-CSF-encoding gene (Csf2) and cooperates with c-Rel in the transactivation of this target gene. PMID: 24899500
- GM-SCF, IL-21 and Rae1 expression, alone or in combination, induces a cellular immune response against H22 tumor cells. PMID: 24350772
- Data indicate that GM-CSF knockout mice exhibit a unique mix of M1-M2 macrophage phenotypes inalveolar macrophages. PMID: 24044676
顯示更多
收起更多
-
亞細(xì)胞定位:Secreted.
-
蛋白家族:GM-CSF family
-
數(shù)據(jù)庫(kù)鏈接:
Most popular with customers
-
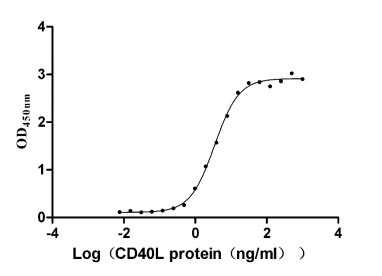
Recombinant Human Tumor necrosis factor receptor superfamily member 5 (CD40), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
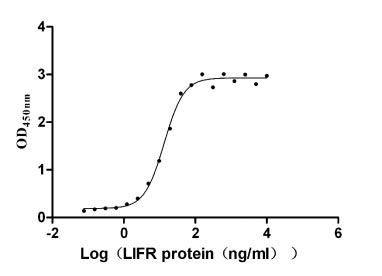
Recombinant Human Leukemia inhibitory factor receptor (LIFR), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
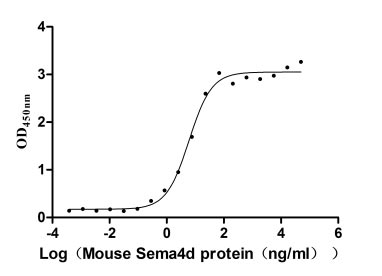
Recombinant Mouse Semaphorin-4D (Sema4d), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
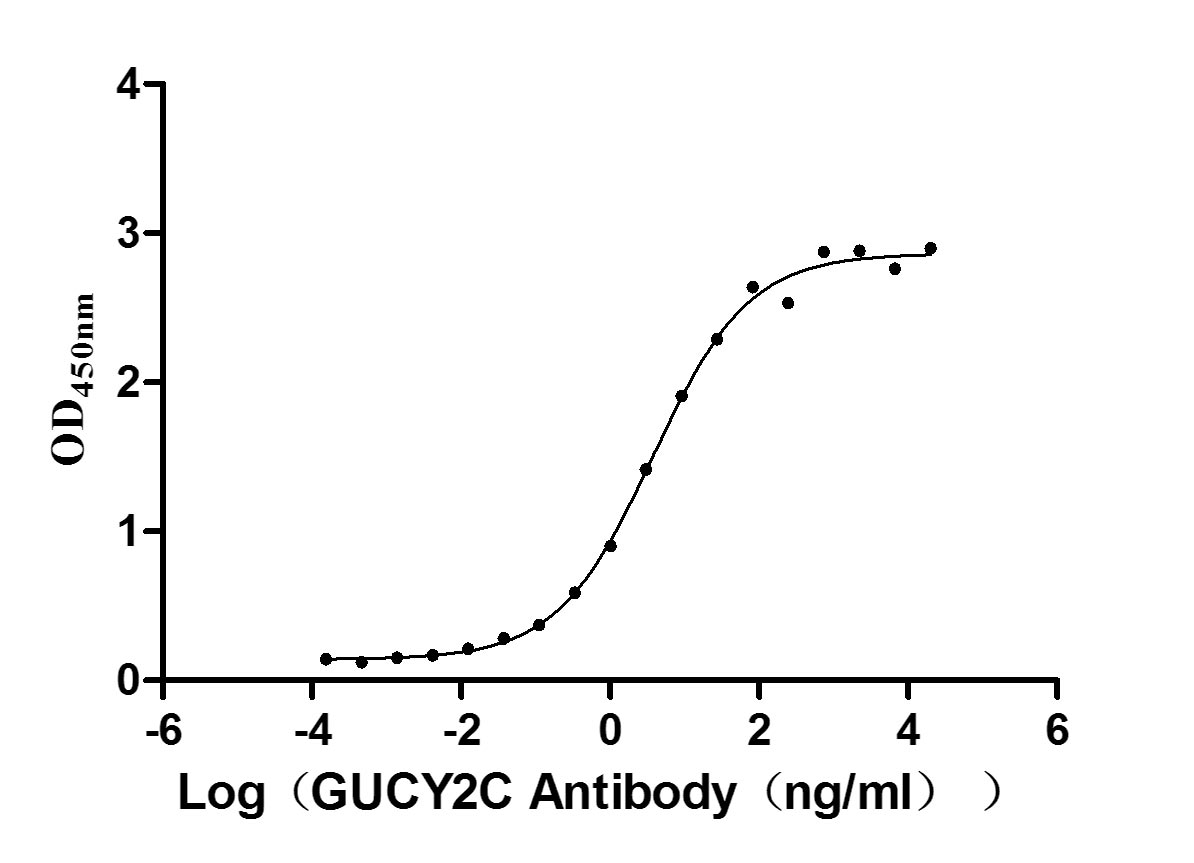
Recombinant Human Heat-stable enterotoxin receptor (GUCY2C), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
-AC1.jpg)
Recombinant Human C-X-C chemokine receptor type 4 (CXCR4)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Macaca fascicularis CD93 molecule (CD93), partial (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
-AC1.jpg)
Recombinant Human Interleukin-17A (IL17A) (T26A) (Active)
Express system: Baculovirus
Species: Homo sapiens (Human)
-
Express system: Mammalian cell
Species: Homo sapiens (Human)









