Recombinant Mouse Frataxin, mitochondrial (Fxn),partial
In Stock-
中文名稱:小鼠Fxn重組蛋白
-
貨號(hào):CSB-YP009086MO
-
說明書:
-
規(guī)格:¥2208
-
圖片:
-
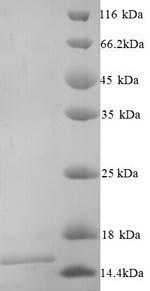 (Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
-
-
其他:
產(chǎn)品詳情
-
純度:Greater than 90% as determined by SDS-PAGE.
-
基因名:
-
Uniprot No.:
-
別名:Fxn; FrdaFrataxin; mitochondrial; Fxn; EC 1.16.3.1) [Cleaved into: Frataxin intermediate form; Frataxin mature form]
-
種屬:Mus musculus (Mouse)
-
蛋白長度:Partial
-
來源:Yeast
-
分子量:16.4kDa
-
表達(dá)區(qū)域:78-207aa
-
氨基酸序列LGTLDNPSSLDETAYERLAEETLDSLAEFFEDLADKPYTLEDYDVSFGDGVLTIKLGGDLGTYVINKQTPNKQIWLSSPSSGPKRYDWTGKNWVYSHDGVSLHELLARELTKALNTKLDLSSLAYSGKGT
Note: The complete sequence may include tag sequence, target protein sequence, linker sequence and extra sequence that is translated with the protein sequence for the purpose(s) of secretion, stability, solubility, etc.
If the exact amino acid sequence of this recombinant protein is critical to your application, please explicitly request the full and complete sequence of this protein before ordering. -
蛋白標(biāo)簽:N-terminal 6xHis-tagged
-
產(chǎn)品提供形式:Liquid or Lyophilized powder Warning: in_array() expects parameter 2 to be array, null given in /www/web/cusabio_cn/public_html/caches/caches_template/default/content/show_product_protein.php on line 662
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
緩沖液:If the delivery form is liquid, the default storage buffer is Tris/PBS-based buffer, 5%-50% glycerol.
Note: If you have any special requirement for the glycerol content, please remark when you place the order.
If the delivery form is lyophilized powder, the buffer before lyophilization is Tris/PBS-based buffer, 6% Trehalose. -
儲(chǔ)存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質(zhì)期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:3-7 business days
-
注意事項(xiàng):Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet & COA:Please contact us to get it.
產(chǎn)品評(píng)價(jià)

相關(guān)產(chǎn)品
靶點(diǎn)詳情
-
功能:Promotes the biosynthesis of heme and assembly and repair of iron-sulfur clusters by delivering Fe(2+) to proteins involved in these pathways. May play a role in the protection against iron-catalyzed oxidative stress through its ability to catalyze the oxidation of Fe(2+) to Fe(3+); the oligomeric form but not the monomeric form has in vitro ferroxidase activity. May be able to store large amounts of iron in the form of a ferrihydrite mineral by oligomerization. Modulates the RNA-binding activity of ACO1.
-
基因功能參考文獻(xiàn):
- This study demonstrated that mouse model of Friedreich's ataxia showed that decreased grip strength endurance time , threshold of peripheral sensitivity using Von Frey monofilaments and gait parameters. PMID: 27575947
- Here, the authors show that loss of Fxn in the nervous system in mice also activates an iron/sphingolipid/PDK1/Mef2 pathway, indicating that the mechanism is evolutionarily conserved. PMID: 27901468
- The in vitro antioxidant treatments trigger the axonal re-growth and the increase in stable MTs in shFxn, thus contributing to identify new neuronal targets of oxidation in this disease and providing a novel approach for antioxidant therapies. PMID: 27516386
- Retinal FXN levels are increased in response to ischemia. Furthermore, elevated FXN levels had a clear neuroprotective effect as shown by increased ganglion cell survival after acute retinal ischemia/reperfusion. Frataxin's neuroprotective effect was associated with an upregulation of antioxidative enzymes. PMID: 27537261
- Frataxin Deficiency Promotes Excess Microglial DNA Damage and Inflammation that Is Rescued by PJ34 PMID: 26954031
- Frataxin-deficient mice, which had higher mitochondrial iron loading, showed impaired airway mucociliary clearance and higher pulmonary inflammation at baseline. PMID: 26752519
- Using a mouse model of hepatic FXN deficiency in combination with mice deficient for IRP1, a key regulator of cellular iron metabolism, we show that IRP1 activation in conditions of Fe-S deficiency increases the available cytosolic labile iron pool PMID: 25651183
- The Fxn KO/Mck mice tested from one to two months of age showed abnormal gait patterns accompanied by a loss in motor skills PMID: 25765157
- Reduced expression of frataxin in Friedreich's ataxia leads to elevation of COX2-mediated oxylipin synthesis stimulated by increases in transcription factors that respond to increased reactive oxygen species. PMID: 25104852
- Frataxin-deficient cells showed a specific inhibition of mitochondrial Complex I activity already at 70% residual frataxin levels, whereas the glutathione imbalance progressively increased after silencing. PMID: 24714088
- The results support a mechanistic hypothesis in which frataxin deficiency decreases Nrf2 expression in vivo, causing the sensitivity to oxidative stress in target tissues the DRG and the cerebella, which contributes to the process of neurodegeneration. PMID: 23350650
- rescue of the Friedreich ataxia knockout mutation in transgenic mice containing an FXN-EGFP genomic reporter PMID: 24667739
- these results indicate that IGF-I exerts cell-context neuroprotection in frataxin deficiency that maybe therapeutically effective. PMID: 23039828
- Data show that the respiratory chain defects accompanying frataxin deficiency cause progressive hyperacetylation of cardiac mitochondrial proteins due to the inhibition of SIRT3 deacetylase. PMID: 22394676
- dual, pro-proliferative but chemosensitizing role in astrocytic tumors PMID: 21863062
- Distribution of frataxin in eye retina of normal mice and of transgenic R7E mice with retinal degeneration PMID: 20799613
- As an attempt to generate a mouse model of Friedreich ataxia, we introduced a (GAA)(230) repeat within the mouse frataxin gene by homologous recombination. PMID: 11852098
- studies indicate an association between frataxin deficiency, iron deposits and cardiac fibrosis, but no obvious association between iron accumulation and neurodegeneration similar to Friedreich ataxia could be detected PMID: 12880182
- frataxin has a role in cellular growth arrest and apoptosis, and its absence can cause experimental diabetes PMID: 12925693
- We have generated two mouse models for FRDA that specifically develop progressive mixed cerebellar and sensory ataxia, the most prominent neurological features of Friedreich ataxia with a slowly progressive neurological degeneration PMID: 14985441
- frataxin may act as a mitochondrial tumor suppressor protein PMID: 16278235
- The identification of a core set of genes changing early in the FRDA pathogenesis can be a useful tool in both clarifying the disease process and in evaluating new therapeutic strategies. PMID: 16442805
- Reduced expression of mitochondrial frataxin in mice exacerbates diet-induced obesity. PMID: 17404227
- The fxn GAA repeat expansion mutation induces comparable epigenetic changes in human and transgenic mouse brain and heart. PMID: 18045775
- loss of Fxn markedly alters cellular Fe trafficking and Fe chelation limits myocardial hypertrophy in the mutant PMID: 18621680
- Frataxin deficiency causes upregulation of mitochondrial Lon and ClpP proteases and severe loss of mitochondrial Fe-S proteins. PMID: 19154341
- The PPARG pathway is dysregulated in frataxin deficient mice and Friedreich's ataxia patients. PMID: 19376812
- We showed that complete absence of murine frataxin in fibroblasts inhibits cell division and leads to cell death. This lethal phenotype was rescued through transgenic expression of human wild type as well as mutant (hFXN(G130V) and hFXN(I154F)) frataxin PMID: 19629184
- Results enable the construction of a model explaining the cytosolic iron deficiency and mitochondrial iron loading in the absence of frataxin, which is important for understanding the pathogenesis of Friedreich's ataxia. PMID: 19805308
顯示更多
收起更多
-
亞細(xì)胞定位:Cytoplasm, cytosol. Mitochondrion. Note=PubMed:17597094 describes localization exclusively in mitochondria.
-
蛋白家族:Frataxin family
-
組織特異性:Heart, liver, skeletal muscle, kidney, spleen and thymus. Weakly expressed in the brain and lung.
-
數(shù)據(jù)庫鏈接:
Most popular with customers
-

Recombinant Severe acute respiratory syndrome coronavirus Spike glycoprotein (S), partial (Active)
Express system: Mammalian cell
Species: Human SARS coronavirus (SARS-CoV) (Severe acute respiratory syndrome coronavirus)
-
Recombinant Human Tumor necrosis factor receptor superfamily member 11B (TNFRSF11B) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Receptor tyrosine-protein kinase erbB-3 (ERBB3), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
-AC1.jpg)
Recombinant Human T-cell surface protein tactile (CD96), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-

Recombinant Human Carcinoembryonic antigen-related cell adhesion molecule 6 (CEACAM6) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Rat Gastric inhibitory polypeptide receptor (Gipr), partial (Active)
Express system: Mammalian cell
Species: Rattus norvegicus (Rat)
-
Recombinant Human Cytotoxic and regulatory T-cell molecule (CRTAM), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)









