Recombinant Human E3 ubiquitin-protein ligase UHRF1 (UHRF1)
-
中文名稱:人UHRF1重組蛋白
-
貨號:CSB-EP847229HU-B
-
說明書:
-
規格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
產品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:UHRF1
-
Uniprot No.:
-
別名:Ac2-121; AL022808; E3 ubiquitin-protein ligase UHRF1; EC 6.3.2.-; FLJ21925; hNP95; hUHRF1; HuNp95; ICBP90; Inverted CCAAT box binding protein of 90 kDa; Inverted CCAAT box binding protein, 90-kD; Inverted CCAAT box-binding protein of 90 kDa; Liver regeneration-related protein LRRG126; MGC138707; NP95; Nuclear phosphoprotein 95; Nuclear phosphoprotein, 95-KD; Nuclear protein 95; Nuclear zinc finger protein Np95; RING finger protein 106; RNF106; TDRD22; Transcription factor ICBP90; Ubiquitin like containing PHD and RING finger domains protein 1; Ubiquitin like PHD and RING finger domain containing protein 1; Ubiquitin-like PHD and RING finger domain-containing protein 1; Ubiquitin-like protein containing PHD and RING finger domains 1; Ubiquitin-like with PHD and ring finger domains 1; Ubiquitin-like, containing PHD and RING finger domains, 1; Ubiquitin-like-containing PHD and RING finger domains protein 1; UHRF1; UHRF1_HUMAN
-
種屬:Homo sapiens (Human)
-
蛋白長度:full length protein
-
表達區域:1-793
-
氨基酸序列MWIQVRTMDG RQTHTVDSLS RLTKVEELRR KIQELFHVEP GLQRLFYRGK QMEDGHTLFD YEVRLNDTIQ LLVRQSLVLP HSTKERDSEL SDTDSGCCLG QSESDKSSTH GEAAAETDSR PADEDMWDET ELGLYKVNEY VDARDTNMGA WFEAQVVRVT RKAPSRDEPC SSTSRPALEE DVIYHVKYDD YPENGVVQMN SRDVRARART IIKWQDLEVG QVVMLNYNPD NPKERGFWYD AEISRKRETR TARELYANVV LGDDSLNDCR IIFVDEVFKI ERPGEGSPMV DNPMRRKSGP SCKHCKDDVN RLCRVCACHL CGGRQDPDKQ LMCDECDMAF HIYCLDPPLS SVPSEDEWYC PECRNDASEV VLAGERLRES KKKAKMASAT SSSQRDWGKG MACVGRTKEC TIVPSNHYGP IPGIPVGTMW RFRVQVSESG VHRPHVAGIH GRSNDGAYSL VLAGGYEDDV DHGNFFTYTG SGGRDLSGNK RTAEQSCDQK LTNTNRALAL NCFAPINDQE GAEAKDWRSG KPVRVVRNVK GGKNSKYAPA EGNRYDGIYK VVKYWPEKGK SGFLVWRYLL RRDDDEPGPW TKEGKDRIKK LGLTMQYPEG YLEALANRER EKENSKREEE EQQEGGFASP RTGKGKWKRK SAGGGPSRAG SPRRTSKKTK VEPYSLTAQQ SSLIREDKSN AKLWNEVLAS LKDRPASGSP FQLFLSKVEE TFQCICCQEL VFRPITTVCQ HNVCKDCLDR SFRAQVFSCP ACRYDLGRSY AMQVNQPLQT VLNQLFPGYG NGR
-
蛋白標簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產品評價

相關產品
靶點詳情
-
功能:Multidomain protein that acts as a key epigenetic regulator by bridging DNA methylation and chromatin modification. Specifically recognizes and binds hemimethylated DNA at replication forks via its YDG domain and recruits DNMT1 methyltransferase to ensure faithful propagation of the DNA methylation patterns through DNA replication. In addition to its role in maintenance of DNA methylation, also plays a key role in chromatin modification: through its tudor-like regions and PHD-type zinc fingers, specifically recognizes and binds histone H3 trimethylated at 'Lys-9' (H3K9me3) and unmethylated at 'Arg-2' (H3R2me0), respectively, and recruits chromatin proteins. Enriched in pericentric heterochromatin where it recruits different chromatin modifiers required for this chromatin replication. Also localizes to euchromatic regions where it negatively regulates transcription possibly by impacting DNA methylation and histone modifications. Has E3 ubiquitin-protein ligase activity by mediating the ubiquitination of target proteins such as histone H3 and PML. It is still unclear how E3 ubiquitin-protein ligase activity is related to its role in chromatin in vivo. Plays a role in DNA repair by cooperating with UHRF1 to ensure recruitment of FANCD2 to interstrand cross-links (ICLs) leading to FANCD2 activation.
-
基因功能參考文獻:
- Our novel findings strongly indicate that inactivation of the Notch signaling pathway impedes hepatocellular carcinoma progress via reduction of ICBP90. PMID: 30309512
- Using an integrative approach that combines small angle X-ray scattering, NMR spectroscopy, and molecular dynamics simulations, we characterized the dynamics of the tandem tudor domain-plant homeodomain (TTD-PHD) histone reader module, including its 20-residue interdomain linker. PMID: 29074623
- UHRF1 deficiency leads to the induction of EMT by activating the CXCR4/AKT-JNK/IL-6 signaling pathway, thereby contributing to the expansion of cancer stem-like cells PMID: 28584306
- studies support a noncompetitive model for UHRF1 and DNMT1 chromatin recruitment to replicating chromatin and define a role for hemimethylated linker DNA as a regulator of UHRF1 ubiquitin ligase substrate selectivity. PMID: 30104358
- Lys4 methylation on H3 peptide has an insignificant effect on combinatorial recognition of R2 and K9me2 on H3 by the UHRF1 TTD-PHD. We propose that subtle variations of key residues at the binding pocket determine status specific recognition of histone methyl-lysines by the reader domains. PMID: 29656054
- Study provides evidence that elevated expression of UHRF1 plays an important role in melanoma cell proliferation and progression, and it can be used as a prognostic biomarker for melanoma. PMID: 29620240
- The emodininduced downregulation of UHRF1 led to an increase in the level of DNA methyltransferase 3A. PMID: 28901428
- Findings confirmed that UHRF1 is a gene driver in medulloblastoma (MB) and, revealed that UHRF1 is in a modulation axis downstream of miR-378. Its promoter region is targeted by miR-378 which negatively regulates its expression in MB cells. PMID: 28901497
- UHRF1 controls both CDH1 and antisense-directed non coding RNA RCDH1-AS by directly binding this bidirectional promoter region. PMID: 29466696
- Epigenetic enhancement of the post-replicative DNA mismatch repair of mammalian genomes by a Hemi-(m)CpG-Np95-Dnmt1 axis has been demonstrated for humans and mice. PMID: 27886214
- These results suggest that tumor-promoting functions of UHRF1 in retinoblastoma are largely independent of its role in DNA methylation. PMID: 28467809
- UHRF1 is a key epigenetic regulator of DNA methylation in esophageal squamous cell carcinoma and might be a potential target for cancer treatment PMID: 27507047
- Elevated UHRF1 expression contributes to poor prognosis by promoting cell proliferation and metastasis in hepatocellular carcinoma. PMID: 28060737
- Results find the overexpression of the anti-apoptotic UHRF1 to coordinate the epigenetic silencing of several tumor suppressor genes in many human haematological and solid cancers causing apoptosis inhibition. [review] PMID: 27839516
- our present data demonstrated that both strands of miR-145 (miR-145-5p: guide-strand and miR-145-3p: passenger-strand) play pivotal roles in bladder cancer cells by regulating UHRF1 PMID: 27072587
- The results indicated that globular adiponectin inhibited the prooncogenic effects of leptin via AdipoR2-mediated suppression of UHRF1 PMID: 28285359
- Hemi-methylated CpGs DNA recognition activates UHRF1 ubiquitylation towards multiple lysines on the H3 tail adjacent to the UHRF1 histone-binding site. PMID: 27595565
- Down-regulation of UHRF1 is an important mechanism of PIM1-mediated cellular senescence. PMID: 28394343
- UHRF1 gene expression levels correlates with the major pathological characteristics of transitional cell carcinoma samples and with the clinical outcome of those patients. Determination of UHRF1 gene expression could have a potential to be used as a sensitive molecular marker in patients with urinary bladder cancer. PMID: 28128913
- decreased UHRF1 expression is a key initial event in the suppression of DNMT1-mediated DNA methylation and in the consequent induction of senescence via increasing WNT5A expression PMID: 28100769
- Our results provide insights into the molecular basis of DNMT1 dysfunction in hereditary sensory and autonomic neuropathies with dementia and hearing loss patients and emphasize the importance of the TS domain in the regulation of DNA methylation in pluripotent and differentiating cells PMID: 28334952
- results identify UHRF1 as a novel HSP90 client protein and shed light on the regulation of UHRF1 stability and function. PMID: 27489107
- Down-regulation of UHRF1 by RNAi inhibited proliferation and clonogenic ability of medullobastoma cell lines with cell cycle arrest in G1/G2-phase. Cells transfected with lenti-shUHRF1 showed increased p16 expression and location shift of CDK4 in medulloblastoma cells. PMID: 27449774
- Our study uncovered that UHRF1 overexpressed in cervical squamous cell carcinoma and silent UHRF1 gene can reduce tumorigenicity of the CaSki cells by decreasing its proliferation capacity and making it stagnate at G0 and G1 phases as well as accelerating its apoptosis PMID: 27431502
- The overexpression of UHRF1 was correlated with the stage and grade of gastric cancer and is associated with the genotype distribution of MiR-146a rs2910164. PMID: 26896831
- Studies indicate that ubiquitin like with PHD and ring finger domains 1 protein (UHRF1) interacts with DNA interstrand crosslinks (ICLs) both in vitro and in vivo. PMID: 26700964
- Results showed that UHRF1 was overexpressed in almost all of the prostate cancer cell (PCa) lines. Its expression levels were correlated with some clinical features of PCa suggesting it as an independent prognostic factor for biochemical recurrence. PMID: 26884069
- UHRF1 promotes osteosarcoma cell invasion by downregulating the expression of Ecadherin and increasing epithelialtomesenchymal transition in an Rb1dependent manner. PMID: 26548607
- Hemi-methylated DNA opens a closed conformation of UHRF1 to facilitate its H3 histone recognition. PMID: 27045799
- UHRF1 was over-expressed in hepatocellular carcinoma tissues compared to the adjacent normal tissues and UHRF1 expression shared significant relevance with distant metastasis, cancer area and HBV. PMID: 26464697
- results suggest that UPAT and UHRF1 may be promising molecular targets for the therapy of colon cancer PMID: 26768845
- These findings identify ICBP90 as a key regulator of MIF transcription and provide functional insight into the regulation of the polymorphic MIF locus. PMID: 26752645
- miR-124 could control the fate of target gene UHRF1 mRNA by binding 3'-UTR PMID: 26310391
- This study showed that UHRF1 interacts directly with BRCA1 and is involved in DNA double-strand break repair. PMID: 26727879
- Findings demonstrated the inhibitory effect of MEG3 in vivo and vitro and illuminated that MEG3 could be a potential biomarker for the survival of hepatocellular carcinoma (HCC) patients. Its expression seemed to be regulated by UHRF1 in HCC. PMID: 25641194
- K-Ras drives UHRF1 expression, establishing a novel link between this oncogene and Nrf2-mediated cellular protection. PMID: 26497117
- UHRF1 is overexpressed in serum and gastric tissue of stomach cancer patients. PMID: 26161699
- The downregulation of UHRF1 by Torin-1 was partially due to a decrease in the UHRF1 mRNA level. Torin-2 effectively inhibited hepatocellular carcinoma cell proliferation through induction of autophagy PMID: 26239364
- UHRF1 promotes proliferation of gastric cancer via mediating tumor suppressor gene hypermethylation. PMID: 26147747
- UHRF1 could act as a new oncogenic factor in ovarian cancer and be a potential molecular target for ovarian cancer gene therapy. PMID: 26070868
- UHRF1 promotes recruitment of lesion-processing activities via its affinity to recognize DNA damage PMID: 25818288
- describe a mechanism of interstrand crosslink (ICL) sensing and propose that UHRF1 is a critical factor that binds to ICLs. In turn, this binding is necessary for the subsequent recruitment of FANCD2, which allows the DNA repair process to initiate PMID: 25801034
- results are consistent with a "reader" role, in which the SRA domain scans DNA sequences for hemimethylated CpG sites without perturbation of the structure of contacted nucleotides. PMID: 26368281
- High UHRF1 expression is associated with ovarian cancer. PMID: 25743796
- The global DNA hypomethylation induced by the DNMT1/PCNA/UHRF1 disruption is an oncogenic event of human tumorigenesis. PMID: 24637615
- miR-9 could repress the expression of UHRF1, and function as a tumor-suppressive microRNA in colorectal cancer. PMID: 25940709
- UHRF1 may play a pivotal role in suppressing the malignant alteration of cancer cells. PMID: 25572953
- Promoter hypermethylation and down-expression of let-7a-3 may play a role in DN by targeting UHRF1. PMID: 26049093
- UHRF1 regulates p53 ubiquitination and p53-dependent cell apoptosis in clear cell renal cell carcinoma. PMID: 26102039
- our study demonstrated that UHRF1 played an oncogenic role in the progression of pancreatic cancer, and UHRF1 might be a promising target for the treatment of pancreatic cancer. PMID: 25416862
顯示更多
收起更多
-
相關疾病:Defects in UHRF1 may be a cause of cancers. Overexpressed in many different forms of human cancers, including bladder, breast, cervical, colorectal and prostate cancers, as well as pancreatic adenocarcinomas, rhabdomyosarcomas and gliomas. Plays an important role in the correlation of histone modification and gene silencing in cancer progression. Expression is associated with a poor prognosis in patients with various cancers, suggesting that it participates in cancer progression.
-
亞細胞定位:Nucleus. Note=Localizes to replication foci. Enriched in pericentric heterochromatin. Also localizes to euchromatic regions.
-
組織特異性:Expressed in thymus, bone marrow, testis, lung and heart. Overexpressed in breast cancer.
-
數據庫鏈接:
Most popular with customers
-
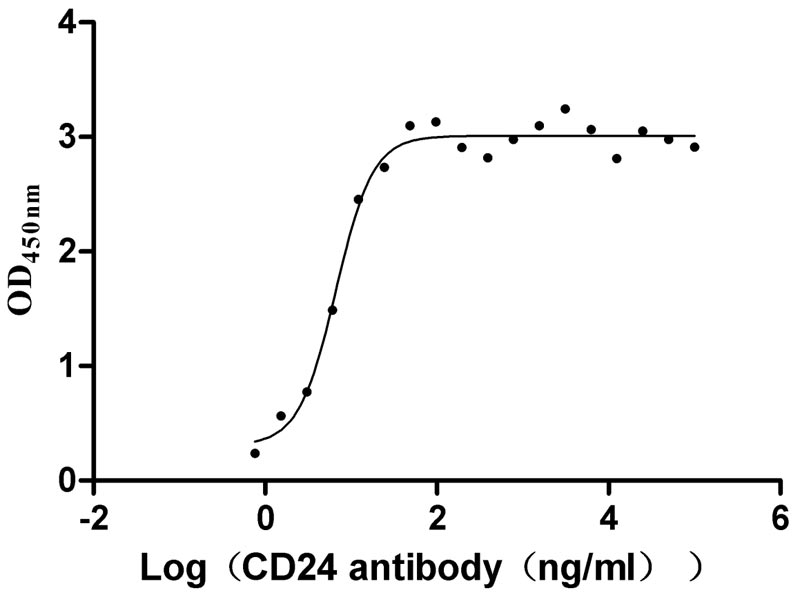
Recombinant Human Signal transducer CD24 (CD24)-Nanoparticle (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
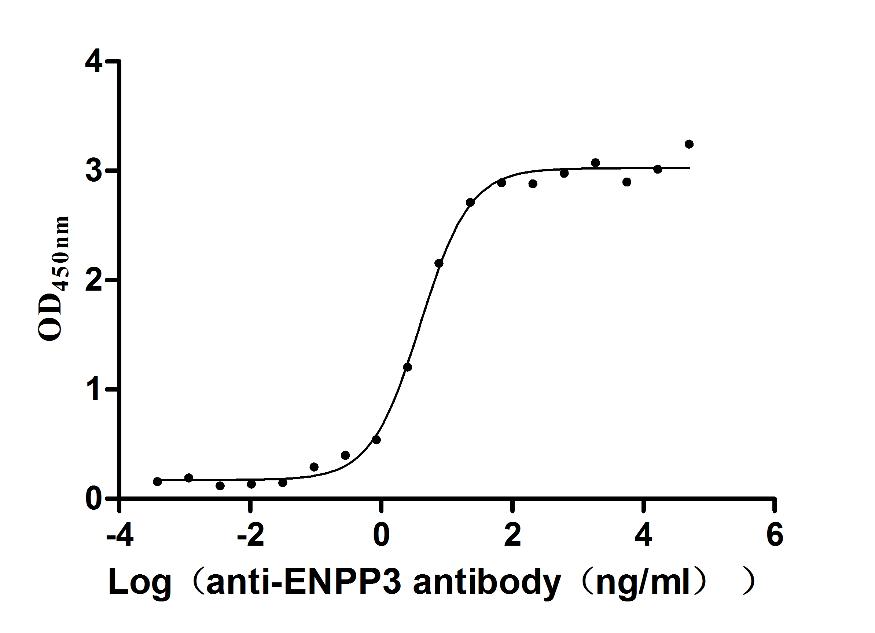
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
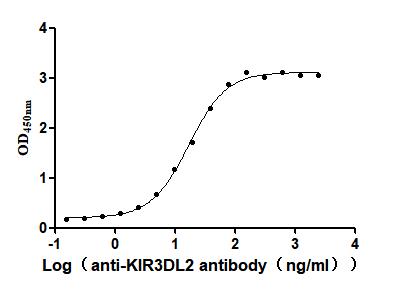
Recombinant Human Killer cell immunoglobulin-like receptor 3DL2 (KIR3DL2), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
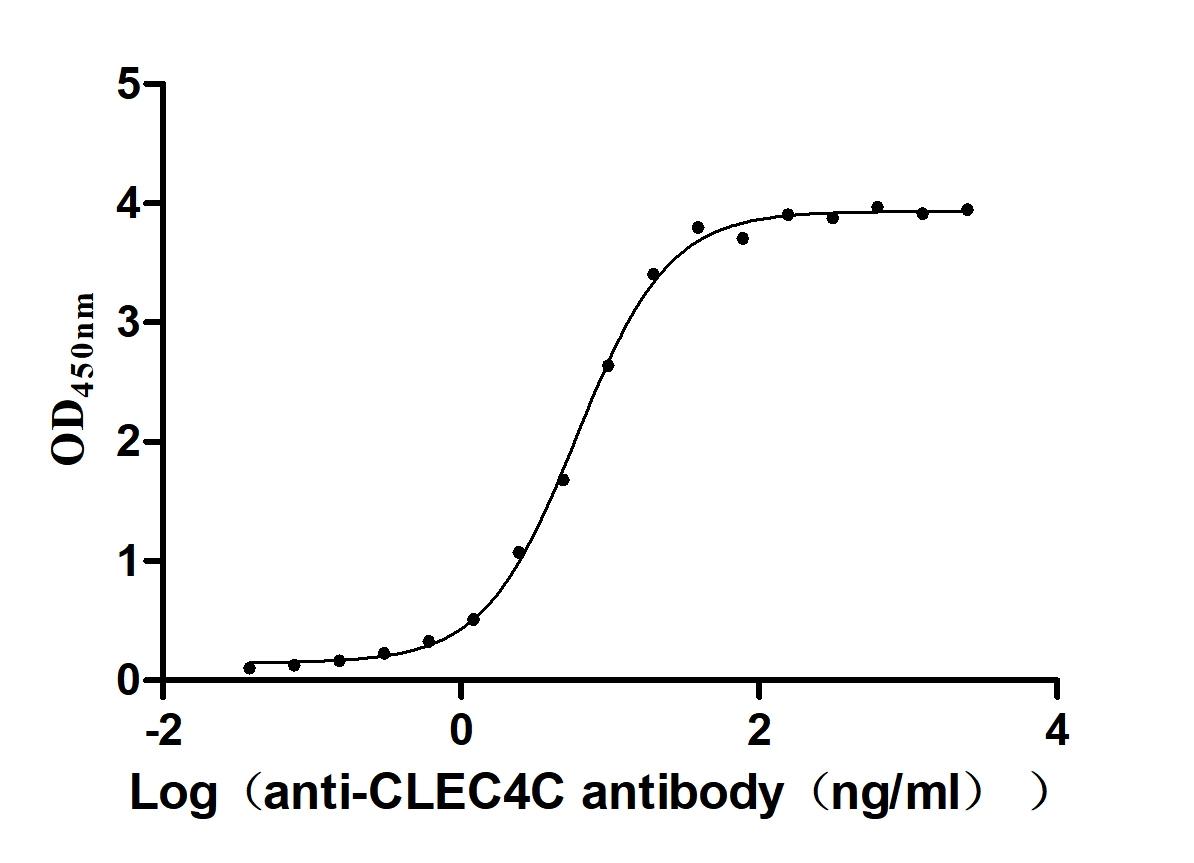
Recombinant Macaca fascicularis C-type lectin domain family 4 member C(CLEC4C), partial (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
Recombinant Human B- and T-lymphocyte attenuator(BTLA), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Macaca fascicularis Zinc transporter ZIP6 isoform X1(SLC39A6),partial (Active)
Express system: Baculovirus
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
-AC1.jpg)
Recombinant Human Tumor necrosis factor ligand superfamily member 15(TNFSF15) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)









