Mouse Interleukin 4,IL-4 ELISA KIT
-
中文名稱:小鼠白介素4(IL-4)酶聯(lián)免疫試劑盒
-
貨號(hào):CSB-E04634m
-
規(guī)格:96T/48T
-
價(jià)格:¥3200/¥2500
-
其他:

產(chǎn)品詳情
-
產(chǎn)品描述:小鼠白介素4(IL-4)酶聯(lián)免疫試劑盒(CSB-E04634m)為雙抗夾心法ELISA試劑盒,定量檢測(cè)血清、血漿、組織勻漿樣本中的IL4含量。IL4是重要的細(xì)胞因子,在免疫調(diào)節(jié)等方面有重要作用。它可調(diào)控Th2細(xì)胞分化,參與過(guò)敏反應(yīng)等。研究機(jī)制上圍繞其信號(hào)通路展開,明確其與受體結(jié)合及下游信號(hào)激活過(guò)程,有望為免疫相關(guān)疾病如哮喘、特應(yīng)性皮炎等治療提供靶點(diǎn)。試劑盒檢測(cè)范圍為31.25 pg/ml-2000 pg/ml.,靈敏度為7.8 pg/ml。本試劑盒適用于基礎(chǔ)免疫學(xué)研究、疾病動(dòng)物模型構(gòu)建(如哮喘、特應(yīng)性皮炎等Th2型疾病)、藥物干預(yù)效果評(píng)估等科研場(chǎng)景,為探究IL - 4在免疫應(yīng)答中的調(diào)控機(jī)制及炎癥相關(guān)疾病研究提供可靠工具。本品僅用于科研,不用于臨床診斷,產(chǎn)品具體參數(shù)及操作步驟詳見產(chǎn)品說(shuō)明書。
-
別名:Il4 ELISA kit; Il-4Interleukin-4 ELISA kit; IL-4 ELISA kit; B-cell IgG differentiation factor ELISA kit; B-cell growth factor 1 ELISA kit; B-cell stimulatory factor 1 ELISA kit; BSF-1 ELISA kit; IGG1 induction factor ELISA kit; Lymphocyte stimulatory factor 1 ELISA kit
-
縮寫:
-
Uniprot No.:
-
種屬:Mus musculus (Mouse)
-
樣本類型:serum, plasma, tissue homogenates
-
檢測(cè)范圍:31.25 pg/ml-2000 pg/ml.
-
靈敏度:7.8 pg/ml
-
反應(yīng)時(shí)間:1-5h
-
樣本體積:50-100ul
-
檢測(cè)波長(zhǎng):450 nm
-
研究領(lǐng)域:Immunology
-
測(cè)定原理:quantitative
-
測(cè)定方法:Sandwich
-
精密度:
Intra-assay Precision (Precision within an assay): CV%<8%
Three samples of known concentration were tested twenty times on one plate to assess.
Inter-assay Precision (Precision between assays): CV%<10%
Three samples of known concentration were tested in twenty assays to assess.
-
線性度:
To assess the linearity of the assay, samples were spiked with high concentrations of mouse IL-4 in various matrices and diluted with the Sample Diluent to produce samples with values within the dynamic range of the assay.
Sample
Serum(n=4)
1:1
Average %
88
Range %
80-97
1:2
Average %
97
Range %
90-105
1:4
Average %
94
Range %
92-96
1:8
Average %
90
Range %
86-95
-
回收率:
The recovery of mouse IL-4 spiked to levels throughout the range of the assay in various matrices was evaluated. Samples were diluted prior to assay as directed in the Sample Preparation section.
Sample Type
Average % Recovery
Range
Serum (n=5)
88
83-98
EDTA plasma (n=4)
90
85-97
-
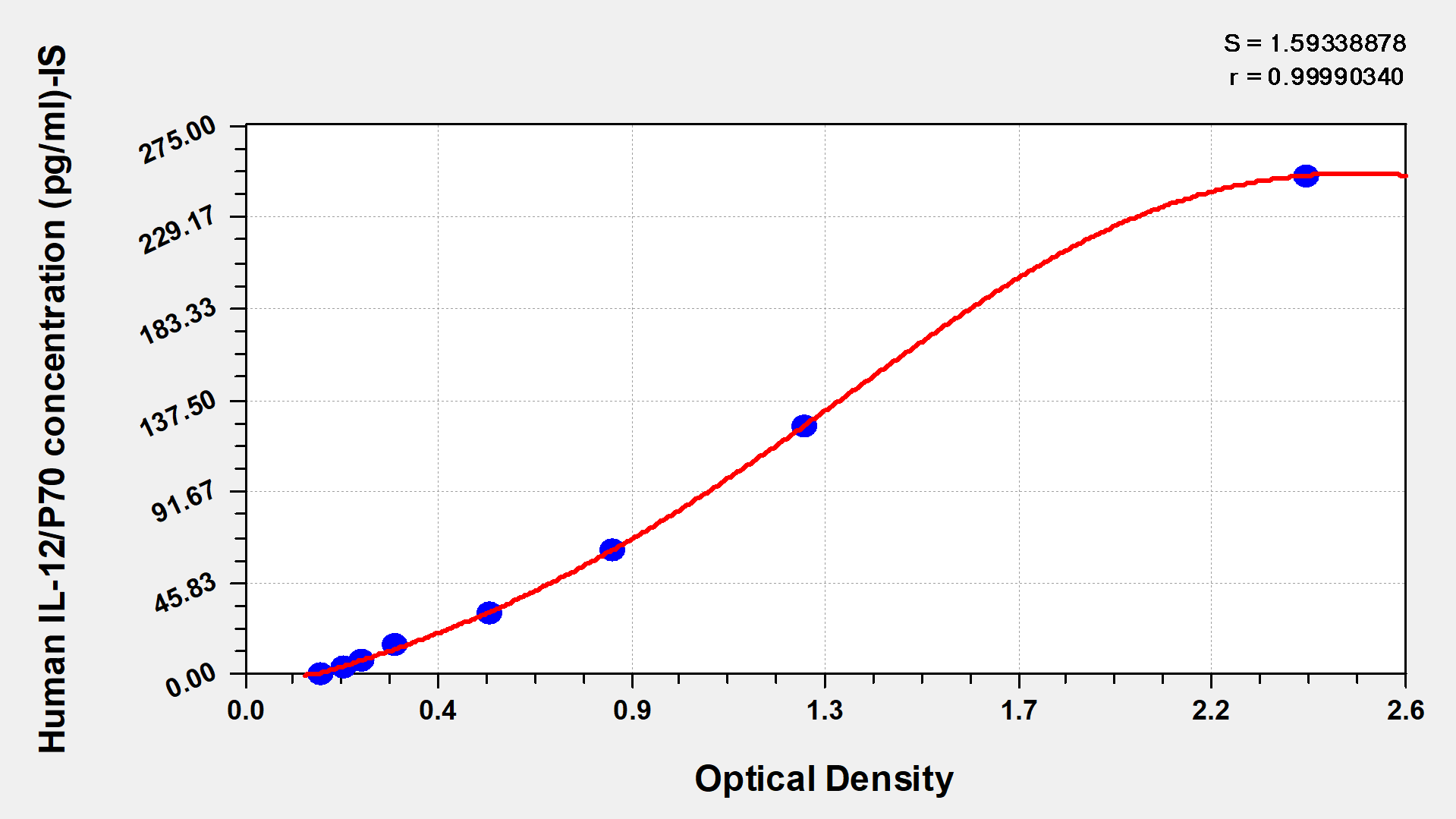
標(biāo)準(zhǔn)曲線:
These standard curves are provided for demonstration only. A standard curve should be generated for each set of samples assayed.

pg/ml
OD1
OD2
Average
Corrected
2000
2.615
2.634
2.625
2.463
1000
1.764
1.697
1.730
1.568
500
1.311
1.228
1.270
1.107
250
0.824
0.829
0.826
0.664
125
0.581
0.621
0.601
0.439
62.5
0.313
0.303
0.308
0.146
31.25
0.257
0.244
0.250
0.088
0
0.159
0.165
0.162
-
數(shù)據(jù)處理:
-
貨期:3-5 working days
引用文獻(xiàn)
- Dendrobium huoshanense polysaccharide inhibits NSCLC proliferation and immune evasion via FXR1-IL-35 axis signaling pathway X Zhu, G Yin, J Xu, X Tang, F Yu,Journal of Natural Medicines,2025
- B-cell epitope peptide immunotherapy alleviates chitin-binding protein-induced type 2 airway inflammation in a Blomia tropicalis-murine model J Zhang, W Luo, YB Cui, B Sun,Respiratory Research,2025
- Butyrate alleviates food allergy by improving intestinal barrier integrity through suppressing oxidative stress‐mediated Notch signaling J Shi, W Mao, Y Song, Y Wang, L Zhang, Y Xu, H Gu,iMeta,2025
- Pruriceptor-like vagal neurons respond to allergic inflammatory mediators JC Wang,Thesis&Dissertation,2024
- In silicon desinging of RANKL-targeting vaccine for protection of osteoporosis based on the epitope of Denosumab T Wu, B Guan, J Luo, L Li, B Zhang, Z Yang,International Immunopharmacology,2025
- Combining Network Pharmacology and Transcriptomics to Investigate the Mechanisms of Yujiang Paidu Decoction in the Treatment of Chronic Rhinosinusitis with Nasal Polyps Y Li, Y Yin, J Xiong, Z Zhang, L Li, B Zhang,Drug Design, Development and Therapy,2024
- Иммунная составляющая патогенетической терапии атопического дерматита ВД Боковой, МА Десятова, АВ Коротков,/,2024
- Double-strand-break repair protein rad21 homolog/Synaptotagmin-7 alleviates Alzheimer's disease in mice by promoting M2 polarization of microglia C Zhu,Brain research bulletin,2024
- Preventive effects of Bifidobacterium lactis Probio-M8 on ovalbumin-induced food allergy in mice J Shi,Food Science and Human Wellness,2024
- Excitatory amino acid transporter supports inflammatory macrophage responses Z Gan,Science Bulletin,2024
- Low expression of lipoic acid synthase aggravates silica-induced pulmonary fibrosis by inhibiting the differentiation of Tregs in mice S Yan,Antioxidants & redox signaling,2023
- USP25 attenuates anti-GBM nephritis in mice by negative feedback regulation of Th17 cell differentiation R Xu,Renal failure,2024
- Iguratimod prevents renal fibrosis in unilateral ureteral obstruction model mice by suppressing M2 macrophage infiltration and macrophage–myofibroblast transition Y Zhou,Renal failure,2024
- METTL3 mediates SOX5 m6A methylation in bronchial epithelial cells to attenuate Th2 cell differentiation in T2 asthma Z Chen,/,2024
- DPP4 Regulates the Th17/IL-17 Axis and Accelerates Epithelial Mesenchymal Transition to Promote Ovalbumin-Induced Asthma in Female C57BL/6J Mice L Li,/,2023
- Lactobacillus rhamnosus Probio-M9 alleviate OVA-sensitized food allergy through modulating gut microbiota and its metabolism J Shi,Food & function,2023
- Pyranochromones with Anti-Inflammatory Activities in Arthritis from Calophyllum membranaceum W Shen,Journal of natural products,2023
- Targeted modulation of intestinal epithelial regeneration and immune response in ulcerative colitis using dual-targeting bilirubin nanoparticles L Pervizaj-Oruqaj,Theranostics,2024
- Pulmonary flora-modified diesel particulate matter induced lung injury via cGAS signaling pathway M Sun,The Science of the total environment,2023
- Food Bioscience W Cui,/,2023
- Astaxanthin protected against the adverse effects induced by diesel exhaust particulate matter via improving membrane stability and anti-oxidative property T Wang,Journal of hazardous materials,2023
- Resveratrol Improves Paclitaxel-Induced Cognitive Impairment in Mice by Activating SIRT1/PGC-1α Pathway to Regulate Neuronal State and Microglia Cell Polarization X Liu,Drug design, development and therapy,2023
- The Trichinella spiralis-derived antigens alleviate HFD-induced obesity and inflammation in mice M Tong,International immunopharmacology,2023
- Cadmium contributes to atherosclerosis by affecting macrophage polarization Y Wan,Food and chemical toxicology,2023
- Nebulized Mycobacterium vaccae protects against asthma by attenuating the imbalance of IRF4/IRF8 expression in dendritic cells QX Sun,Asian Pacific Journal of Tropical Biomedicine,2022
- Increased soluble HLA in COVID-19 present a disease-related, diverse immunopeptidome associated with T cell immunity A Nelde,/,2022
- Mechanical stimulation improves rotator cuff tendon-bone healing via activating IL-4/JAK/STAT signaling pathway mediated macrophage M2 polarization Y Liu,Journal of Orthopaedic Translation,2022
- Glycyrrhizic acid suppresses atopic dermatitis‐like symptoms by regulating the immune balance DD Hou,Journal of cosmetic dermatology,2022
- Aerosol Inhalation of Heat-Killed Clostridium butyricum CGMCC0313-1 Alleviates Allergic Airway Inflammation in Mice L Li,Journal of Immunology Research,2022
- Paclitaxel induces cognitive impairment via necroptosis, decreased synaptic plasticity and M1 polarisation of microglia M Tang,Pharmaceutical biology,2022
- TLR2-ERK signaling pathway regulates expression of galectin-3 in a murine model of OVA-induced allergic airway inflammation Y Lv,/,2022
- N‐acetylcysteine improves diabetic associated erectile dysfunction in streptozotocin‐induced diabetic mice by inhibiting oxidative stress Z Ma,Journal of cellular and molecular medicine,2022
- Combination with Toll-like receptor 4 (TLR4) agonist reverses GITR agonism mediated M2 polarization of macrophage in Hepatocellular carcinoma C Pan,Oncoimmunology,2022
- Baicalin Attenuates Continuous Activation of β-Catenin Induced by Lipopolysaccharide (LPS) and Depression Complicated by Infertility in Male Rats R Fan,Evidence-based complementary and alternative medicine : eCAM,2022
- The N-Formyl Peptide Receptor 2 (FPR2) Agonist MR-39 Improves Ex Vivo and In Vivo Amyloid Beta (1–42)-Induced Neuroinflammation in Mouse Models of Alzheimer’s Disease Ewa Trojan,Molecular neurobiology,2021
- FSTL1 aggravates OVA-induced inflammatory responses by activating the NLRP3/IL-1β signaling pathway in mice and macrophages Y Wang,Inflammation Research,2021
- Identification of histone acetylation in a murine model of allergic asthma by proteomic analysis Y Ren,Experimental biology and medicine,2020
- Extracellular vesicles derived from Trichinella spiralis prevent colitis by inhibiting M1 macrophage polarization X Gao,,acta tropica,2020
- Anti-Helicobacter pylori activity of acomplex mixture of Lactobacillus paracasei HP7 including the extract of Perilla frutescens var. acuta and Glycyrrhiza glabra HA Lee,Laboratory animal research,2020
- Anti-fatigue effects of pea (Pisum sativum L.) peptides prepared by compound protease T Feng,Journal of Food Science and Technology,2020
- Upregulation of IL-4 signaling contributes to aerobic exercise-induced insulin sensitivity Z Chen,Biochem. Biophys. Res. Commun,2020
- H2S alleviates renal injury and fibrosis in response to unilateral ureteral obstruction by regulating macrophage infiltration via inhibition of NLRP3 signaling Zhou Y, et al,Experimental Cell Research,2019
- Activating Macrophages for Enhanced Osteogenic and Bactericidal performance by Cu Ion Release from Micro/Nano-Topographical Coating on Titanium Substrate Huang Q, et al,Acta Biomaterialia,2019
- Evaluations of Alkyl hydroperoxide reductase B cell antigen epitope as a potential epitope vaccine against Campylobacter jejuni HongqiangLou,SAUDI JOURNAL OF BIOLOGICAL SCIENCES,2019
- Anti-inflammatory Property of Galectin-1 in a Murine Model of Allergic Airway Inflammation Yunxiang Lv,Journal of Immunology Research,2019
- Critical role of Tim-3 mediated autophagy in chronic stress induced immunosuppression Qin A, et al,Cell And Bioscience,2019
- Mast cells contribute to Enterovirus 71 infection-induced pulmonary edema in neonatal mice Yuefei Jin.et al,Laboratory Investigation,2018
- IL-4 mediates the delayed neurobehavioral impairments induced by neonatal hepatitis B vaccination that involves the down-regulation of the IL-4 receptor in the hippocampus XiaoWang.et al,Cytokine,2018
- Clonorchis sinensis cyclophilin A immunization protected mice from CLP-induced sepsis Juan Jiang.et al,International Immunopharmacology,2018
- Comparison of the roles of house dust mite allergens, ovalbumin and lipopolysaccharides in the sensitization of mice to establish a model of severe neutrophilic asthma Aijun Jia.et al,Experimental and Therapeutic Medicine,2017
- Punicalagin, a PTP1B inhibitor, induces M2c phenotype polarization via up-regulation of HO-1 in murine macrophages Xu X.et al,Free Radic Biol Med.,2017
- In vitro effect of 4-pentylphenol and 3-methyl-4-nitrophenol on murine splenic lymphocyte populations and cytokine/granzyme production. Yang L.et al,J Immunotoxicol.,2016
- Paeoniflorin Atttenuates Amyloidogenesis and the Inflammatory Responses in a Transgenic Mouse Model of Alzheimer's Disease. Zhang HR.et al,Neurochem Res.,2015
- Walnut Polyphenol Extract Attenuates Immunotoxicity Induced by 4-Pentylphenol and 3-methyl-4-nitrophenol in Murine Splenic Lymphocyte Yang L.et al,Nutrients.,2016
- Mannosylated and lipid A-incorporating cationic liposomes constituting microneedle arrays as an effective oral mucosal HBV vaccine applicable in the controlled temperature chain. Wang T.et al,Colloids Surf B Biointerfaces.,2015
- Non-cultured dermal-derived mesenchymal cells attenuate sepsis induced by cecal ligation and puncture in mice Wang Y. et al,Sci Rep,2015
- JNK-TLR9 signal pathway mediates allergic airway inflammation through suppressing melatonin biosynthesis Wu HM.et al,J Pineal Res.,2016
- Enhancing immune responses to inactivated porcine parvovirus oil emulsion vaccine by co-inoculating porcine transfer factor in mice Wang RN et al,Vaccine,2012
- Paeoniflorin Atttenuates Amyloidogenesis and the Inflammatory Responses in a Transgenic Mouse Model of Alzheimer´s Disease Hong-Ri Zhang. et al,Neurochemical Research,2015
- Passive transfer of tumor-rerived MDSCs inhibits asthma-related airway inflammation Song C et al,Scand J Immunol,2013
產(chǎn)品評(píng)價(jià)

相關(guān)產(chǎn)品
靶點(diǎn)詳情
-
最新研究進(jìn)展:IL4是一種細(xì)胞因子,主要由T細(xì)胞和嗜堿性粒細(xì)胞產(chǎn)生。它在免疫反應(yīng)中發(fā)揮重要作用,如調(diào)節(jié)B細(xì)胞的增殖和分化,增強(qiáng)抗體的產(chǎn)生等。最近的研究表明,IL4在某些腫瘤的免疫治療中具有重要作用。例如,IL4可以增強(qiáng)CAR-T細(xì)胞對(duì)癌細(xì)胞的殺傷作用,并且可以通過(guò)改變腫瘤微環(huán)境中的免疫細(xì)胞和分子來(lái)抑制腫瘤的生長(zhǎng)和擴(kuò)散。
-
功能:Participates in at least several B-cell activation processes as well as of other cell types. It is a costimulator of DNA-synthesis. It induces the expression of class II MHC molecules on resting B-cells. It enhances both secretion and cell surface expression of IgE and IgG1. It also regulates the expression of the low affinity Fc receptor for IgE (CD23) on both lymphocytes and monocytes. Positively regulates IL31RA expression in macrophages. Stimulates autophagy in dendritic cells by interfering with mTORC1 signaling and through the induction of RUFY4.
-
基因功能參考文獻(xiàn):
- Downregulation of macrophage Irs2 by hyperinsulinemia impairs IL-4-indeuced M2a-subtype macrophage activation in obesity. PMID: 30451856
- Ndfip1 preserves Treg lineage stability and immune homeostasis by preventing the expansion of highly proliferative and metabolically active Treg cells and by preventing pathological secretion of IL-4 from Treg cells PMID: 28580955
- IL-4/ STAT6 signaling needs to be well adjusted to ensure proper development and function of homing Th2 cells. PMID: 29738764
- By establishing that IL-4 is posttranslationally regulated by TRX-promoted reduction of a disulfide bond, our findings highlight a novel regulatory mechanism of the type 2 immune response that is specific to IL-4 over IL-13. PMID: 30104382
- The VEGFR1-mediated signaling suppressed IL-4-induced Arg-1 expression. PMID: 29110610
- The results obtained in the present study suggest that a signaling pathway mediated by FcRg or the FcRg-Syk axis is commonly required for innate basophil IL-4 responses under conditions mimicking encounters with allergen sources. PMID: 26703455
- IL-4Delta2 did not compete with IL-4 for IL-4Ralpha binding and did not interfere with the downstream STAT-6 phosphorylation in T cells. PMID: 28917204
- this study shows that IL4 and IL21 cooperate to induce the high Bcl6 protein level required for germinal center formation PMID: 28875978
- The complex role of IL-4 in autoimmunity and cholangitis. PMID: 27721424
- The results demonstrate that IL-4 can restore insulin sensitivity in adipocytes via mechanisms not associated with induced adipogenesis or de novo formation of lipid depots. PMID: 29738684
- Interleukin 4 (IL-4) signaling prevents Chlamydia trachomatis Infection from Inducing upper genital tract (UGT) pathology. PMID: 28765368
- In the lung, surfactant protein A (SP-A) enhanced interleukin-4 (IL-4)-dependent macrophage proliferation and activation, accelerating parasite clearance and reducing pulmonary injury after infection with a lung-migrating helminth. In the peritoneal cavity and liver, C1q enhancement of type 2 macrophage activation was required for liver repair after bacterial infection. PMID: 28495878
- Data, including data from studies using transgenic mice, suggest that over-expression of IL4 (interleukin 4) in thyroid tissue/cells up-regulates expression of Duox1 (dual oxidase 1), Duoxa1 (dual oxidase maturation factor 1), and Slc26a4 (pendrin) in thyroid tissue/cells; expression of Slc5a5 (sodium-iodide symporter) is down-regulated. PMID: 27599561
- we defined a molecular mechanism for IL-4 downregulation of involucrin in keratinocytes, which may play an important role in the pathogenesis of AD. PMID: 26918372
- In this study, the effect of continuous IL-4 delivery or bioactive implant coating that constitutively releases a protein inhibitor of CCL2 signaling (7ND) on particle induced osteolysis were studied in the murine continuous femoral intramedullary particle infusion model PMID: 27114284
- T follicular helper (Tfh) cells arise in tumor-draining lymph nodes where they produce an abundance of IL4. Deletion of IL4-expressing Tfh cells improves antitumor immunity, delays tumor growth, and reduces the generation of immunosuppressive myeloid cells in the lymph nodes. PMID: 27920023
- Findings suggest that interleukin 4 (IL-4) affects anti-tumor immunity and constitutes an attractive therapeutic target to reduce immune suppression in the tumor microenvironment. PMID: 28733709
- this study shows that environmental IL-4 plays a role in conditioning early thymic progenitors lineage choice, which would impact T cell development PMID: 28893952
- this study shows that eosinophils subvert host resistance to an intracellular pathogen by instigating non-protective IL-4 in CCR2(-/-) mice PMID: 27049063
- findings show that during intestinal helminth infection, IL-4 derived from T follicular helper cells is required for IgE class switching and plasmablast formation PMID: 28533444
- Data suggest that Il4 (usually released from helper T-cells) induces Cox1 in macrophages at post-transcriptional level; activation of Ampk (catalytic subunit Prkaa1) by metformin blocks Il4-dependent induction of Cox1 and blocks macrophage polarization/activation. (Il4 = interleukin-4; Cox1 = cyclooxygenase 1; Ampk = AMP-activated protein kinase) PMID: 28684424
- IL-4 is required for the development of ex-Foxp3 T helper 2 cells. PMID: 28507062
- conclude that a state of haploinsufficiency for the Il4 gene locus is specifically relevant for IL-4-dependent IgE responses to allergens with the amount of IL-4 produced in the hemizygous condition falling close to the threshold required for switching to IgE production PMID: 28115531
- priming of T helper cells by IL-6-deficient antigen-presenting dendritic cells preferentially leads to accumulation of a subset of Follicular helper T cells characterized by high expression of GATA3 and IL-4. PMID: 27474166
- eosinophils drive progression of myocarditis to Inflammatory dilated cardiomyopathy (DCMi), cause severe DCMi when present in large numbers, and mediate this process through IL-4. PMID: 28302646
- These data suggest that although IL-4-stimulated alternatively activated macrophages upregulate fatty acid oxidation, fatty acid oxidation is dispensable for macrophage polarization and high-fat diet-induced metabolic dysfunction. Macrophage fatty acid oxidation likely plays a correlative, rather than causative, role in systemic metabolic dysfunction. PMID: 28223293
- Excessive IL-4 levels in the mesenteric lymph nodes (MLNs) directly inhibited the induction of aiTregs and caused enteropathy. The aiTregs generated in the attenuation of T cell-dependent food allergic enteropathy may function differently than aiTregs induced in a tolerance model. PMID: 28234975
- this study shows that wild-type mice develop an eosinophilic Th2 airway disease in response to Alternaria alternata exposure, whereas IL-4-deficient mice exhibit a primarily neutrophilic response PMID: 27815425
- Study showed that the intraperitoneal administration of the exogenous cytokines IFN-gamma (to promote M1 microglia ) and IL-4 (to promote M2 microglia) can correctly modulate the timing of the M1 to M2 ratio to affect epileptogenesis and to improve cognitive function in pilocarpine-induced status epilepticus. PMID: 27956120
- These findings indicate that IL-4, a canonical Th2 cell cytokine, can sometimes promote rather than impair Th1 cell-type immune responses PMID: 27298446
- Keratinocyte gene expression is critically shaped by IL-4, altering cell fate decisions, which are likely important for the clinical manifestations and pathology of allergic skin disease PMID: 27554818
- Data show that lactic acid in tumor microenvironments inhibited interferon-gamma (IFNgamma) and intert=leukin-4 (IL4) productions from NKT cells by inhibiting mammalian target of rapamycin (mTOR) signaling. PMID: 27995420
- this study shows that IL-4-mediated control of the precursor population affects the development of virus-specific CD8+ T-cell memory PMID: 27457412
- IL-4 secretion by group 2 innate lymphoid cells contributes to the allergic response in food allergy by reducing allergen-specific Treg cell and activating mast cell counts PMID: 27177780
- These studies clearly show a crucial role for IL-4 in the induction of airway hyperresponsiveness following Strongyloides venezuelensis infection and for IL-33/ST2 in maintaining airway hyperresponsiveness and lung Th2 responses. PMID: 27102638
- we used recombinant herpes simplex virus vector S4IL4 that encode mouse il4 gene to evaluate the therapeutic potential of IL-4 in naloxone-precipitation morphine withdrawal (MW). One week after microinjection of the vector S4IL4 into the PAG LacZ or mouse IL-4 immunoreactivity in the vlPAG was visualized. ELISA assay showed that vector S4IL4 into the PAG induced the expression of IL-4 PMID: 28206989
- this study shows that IL-4 is increased in the brain of T cell receptor transgenic mice, which exhibit impaired memory and adult hippocampal neurogenesis PMID: 27432189
- this study shows that il-4 plays an important role in ESAT-6-induced MCP-1 production by macrophages, and suggest a pathway with significance in tuberculosis pathogenesis PMID: 27154637
- indicate that Siglec-9 affects several different signaling pathways in IL-4-stimulated macrophages, which resulted in enhanced induction of Arg1 in Siglec-9-expressing RAW264 cells PMID: 26540411
- Oct-1 and Oct-2 bound within the Il4 promoter region and the Th2 LCR PMID: 26840450
- Loss of IL-4 promoted expression of M1 microglia/macrophage markers and impaired expression of M2 markers after transient or permanent middle cerebral artery occlusion. PMID: 26732561
- These results indicate a positive role of Batf in promoting the generation of pro-allergic IL-4-producing T-follicular helper cells. PMID: 26278622
- IL-4 induces miR-142-5p and downregulates miR-130a-3p in macrophages, regulating macrophage profibrogenic gene expression in chronic inflammation. PMID: 26436920
- these findings underscore the important collaboration between IL-4 and IL-21 in shaping T-dependent B cells antibody responses. PMID: 26491200
- IL-4 KO mice display state, but not trait, anxiety suggesting that reductions in endogenous anti-inflammatory bioactives can engender subtypes of anxiety PMID: 25772794
- physiologic doses of interleukin-4 (IL-4) and interleukin-13 (IL-13) have profound anti-lymphangiogenic effects and potently impair LEC survival, proliferation, migration PMID: 26039103
- Concerted activity of IgG1 antibodies and IL-4/IL-25-dependent effector cells trap helminth larvae in the tissues following vaccination with defined secreted antigens, providing sterile immunity to challenge infection. PMID: 25816012
- may be an important factor in providing 1,25D3-induced protection from experimental autoimmune encephalomyelitis PMID: 25574039
- IL-4-producing DCs are induced under some Th2-provoking situations, and they should play important roles in initiation of Th2 response. PMID: 26363056
- RUN and FYVE domain-containing protein 4 enhances autophagy and lysosome tethering in response to Interleukin-4. PMID: 26416964
顯示更多
收起更多
-
亞細(xì)胞定位:Secreted.
-
蛋白家族:IL-4/IL-13 family
-
數(shù)據(jù)庫(kù)鏈接:
Most popular with customers
-
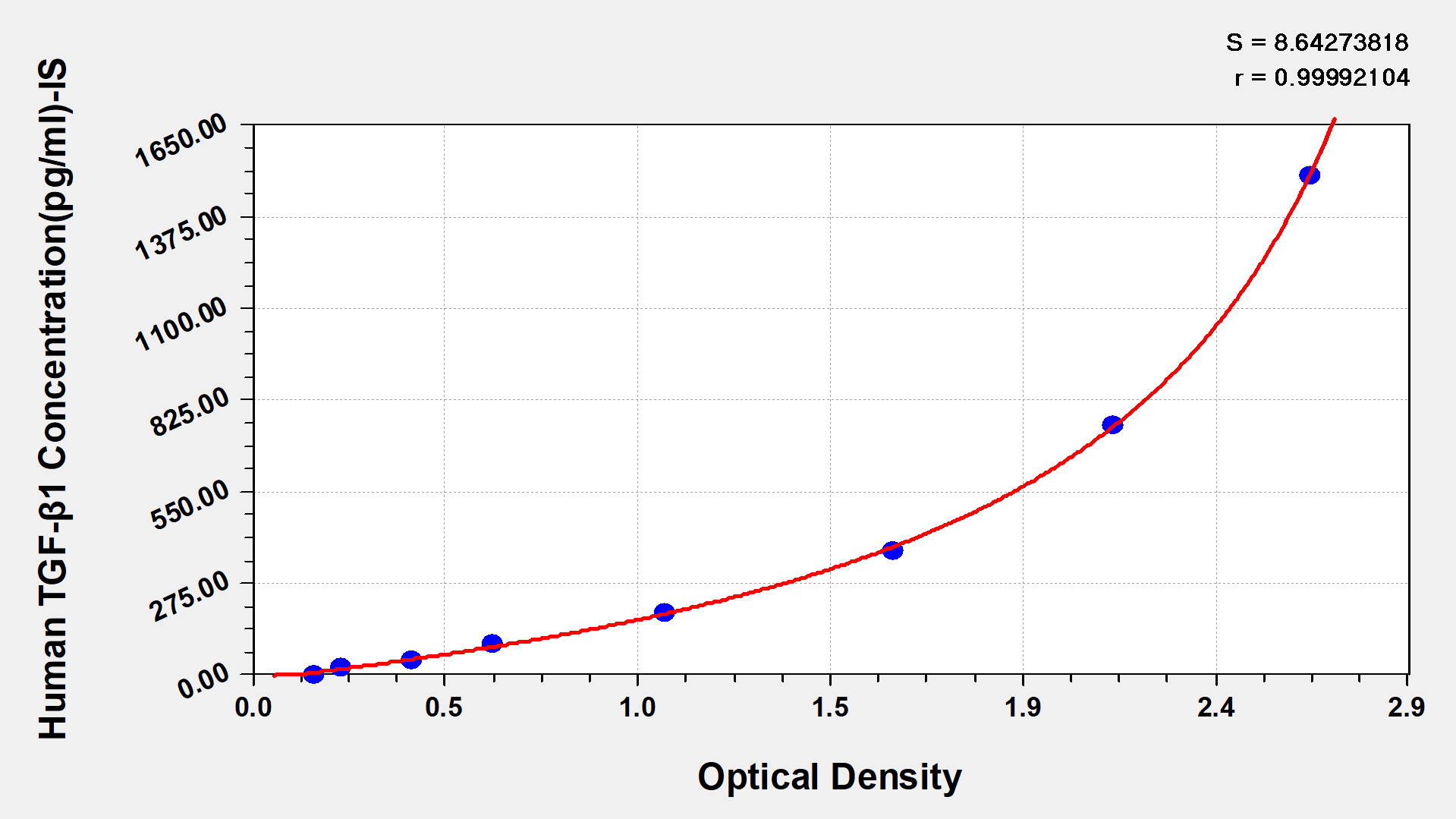
Human Transforming Growth factor β1,TGF-β1 ELISA kit
Detect Range: 23.5 pg/ml-1500 pg/ml
Sensitivity: 5.8 pg/ml
-
-
-
Mouse Tumor necrosis factor α,TNF-α ELISA Kit
Detect Range: 7.8 pg/ml-500 pg/ml
Sensitivity: 1.95 pg/ml
-
-
-
-